

Utilisation de la semence sexée en production bovine
L’utilisation de semence sexée est très développée chez les bovins. Cet article présente différentes facettes de ce sujet : la méthode, ses impacts techniques, sa diffusion et ses conséquences sur les systèmes de production.
Introduction
L’insémination artificielle (IA) s’est développée en France à partir de la fin des années 1940, en particulier chez les bovins (Foote, 2002). Elle consiste à collecter la semence d’un reproducteur mâle, la conditionner, le plus souvent la congeler, puis la transporter et la mettre en place dans les voies génitales de la femelle. Elle permet le découplage entre production de sperme et insémination, ce qui évite le transport des reproducteurs, limite les risques sanitaires, favorise les échanges à plus grande distance et souvent entre pays. L’avantage majeur de l’IA bovine est la dilution de la semence qui permet de produire jusqu’à plusieurs centaines de doses par éjaculat de taureau. Le nombre de descendants par reproducteur peut ainsi être démultiplié, un taureau pouvant produire plusieurs dizaines de milliers de doses en quelques mois. L’impact considérable de l’insémination dans la diffusion du progrès génétique est rappelé par Boichard (2020). Chez les bovins laitiers, un éleveur pratiquant l’IA peut bénéficier d’un progrès génétique de l’ordre de 0,2 à 0,4 écart-type génétique par an, par la simple utilisation de taureaux d’insémination bien choisis, et sans considérer la sélection intratroupeau qu’il peut pratiquer. Le développement de l’IA dépend surtout de son adéquation pratique au système de production. Elle est très majoritaire chez les bovins laitiers, l’éleveur étant proche de ses vaches pour la détection des chaleurs comme pour l’acte d’insémination. Dans les troupeaux de bovins allaitants, elle est limitée par le système de production du fait des difficultés de détection des chaleurs et de contention, les animaux ayant un contact plus limité avec l’homme.
La méiose induisant une ségrégation aléatoire des chromosomes X et Y, on attend en espérance 50 % de spermatozoïdes porteurs du X et 50 % de porteurs du Y. En pratique, on observe à la naissance un peu plus de veaux mâles que de femelles (51 % vs 49 %). Ce léger déséquilibre peut être dû à une survie différentielle de l’embryon lors du développement, ou au poids légèrement plus faible des spermatozoïdes mâles, du fait d’un contenu en ADN légèrement plus faible, les favorisant dans la course pour la fertilisation. La production de semence et sa congélation puis l’insémination artificielle ne modifient pas ce sex-ratio des veaux, avec environ 51 % de mâles parmi les veaux nés de semence conventionnelle. En élevage laitier comme allaitant, où très majoritairement le cheptel femelle est en autorenouvellement, faire naître plus de femelles est pourtant d’intérêt majeur, afin d’accroître notamment les possibilités de sélection intratroupeau. Le conditionnement de la semence pour l’insémination implique différentes procédures de préparation et de contrôle et depuis longtemps, des recherches ont été entreprises pour dévier ce sex-ratio dans la direction désirée. Plusieurs procédés ont été proposés et étudiés (Druart & Ribeiro Bento Dos Santos, 2004), basés sur la différence de quantité d’ADN, de volume ou de masse entre spermatozoïdes X et Y, ou sur des différences d’antigènes de surface. Une équipe japonaise a récemment réussi une séparation des spermatozoïdes X et Y après activation des gènes TLR7/8 localisés sur le chromosome X (Umehara et al., 2020). Mais jusqu’à présent, un seul procédé, basé sur du tri cellulaire, est suffisamment abouti pour une application commerciale à grande échelle. L’objectif de cet article est d’analyser l’intérêt et l’impact du sexage de la semence dans l’espèce bovine et d’en évaluer les conséquences en termes de sélection génétique, de systèmes d’élevage et à l’échelle des filières lait et viande.
1. Technologie appliquée au sexage de la semence
Le principe est de séparer, par un traitement particulier, les spermatozoïdes portant un chromosome X et donnant une femelle, des spermatozoïdes portant un chromosome Y et donnant un mâle. Le procédé appliqué actuellement a été développé par des chercheurs de l’USDA dans les années 1990 (Johnson et al., 1987 ; Johnson & Seidel, 1999) sur différentes espèces de mammifères, puis breveté par la société XY Inc (Johnson, 1992). La société américaine Sexing Technologies a racheté XY en 2007 et propose en exclusivité la prestation de sexage de la semence bovine.
Le procédé repose sur le tri de spermatozoïdes après traitement de la semence avec une substance, le Hoechst 33342, qui se fixe sur l’ADN. Ce produit est fluorescent quand il est éclairé par les UV et les spermatozoïdes traités, éclairés par un laser UV, sont d’autant plus fluorescents qu’ils contiennent plus d’ADN. Le chromosome X étant plus grand que le chromosome Y, les spermatozoïdes femelles ont plus d’ADN (4 %) que les spermatozoïdes mâles et sont légèrement plus fluorescents, et cette différence est utilisée pour séparer les deux types de spermatozoïdes. De façon plus détaillée, la semence est diluée et sa concentration est standardisée, puis elle est traitée au Hoechst 33342 pendant une heure. À l’entrée du trieur de cellules, la solution est finement brumisée de sorte que chaque spermatozoïde est enfermé dans une gouttelette et orienté pour que le signal lumineux soit interprétable, ce qui constitue une des difficultés techniques importantes. En fonction du niveau de fluorescence détecté après éclairage par un laser, les spermatozoïdes sont chargés électriquement positivement ou négativement, et déviés dans l’une ou l’autre direction. Les spermatozoïdes morts, qui n’ont pas intégré de fluorophore Hoechst 33342, sont également éliminés. Les distributions de fluorescence mesurée lors du tri n’étant pas disjointes, on ne peut pas retenir les 50 % de spermatozoïdes du sexe choisi. Pour garantir un niveau de pureté suffisant, on ne retient donc que la partie de la distribution constituée en majorité de spermatozoïdes du sexe choisi. L’objectif fréquemment fixé est une précision de tri de 90 à 95 %. Ces valeurs résultent d’un compromis entre pureté souhaitée et quantités produites, car plus la pureté recherchée est élevée, plus importante est la perte de semence et plus long est le procédé, et donc plus la technique est coûteuse. Le rendement est d’autant meilleur que les deux distributions de fluorescence sont moins chevauchantes et il est important que le signal de fluorescence soit le moins bruité possible. En théorie, le procédé permet de produire des doses mâles ou des doses femelles. Mais compte tenu de la dissymétrie de la sélection pratiquée, la fraction non sélectionnée n’est pas utilisable, car elle ne présente pas un niveau de pureté acceptable pour l’autre sexe. La figure 1 présente le principe de cette séparation.
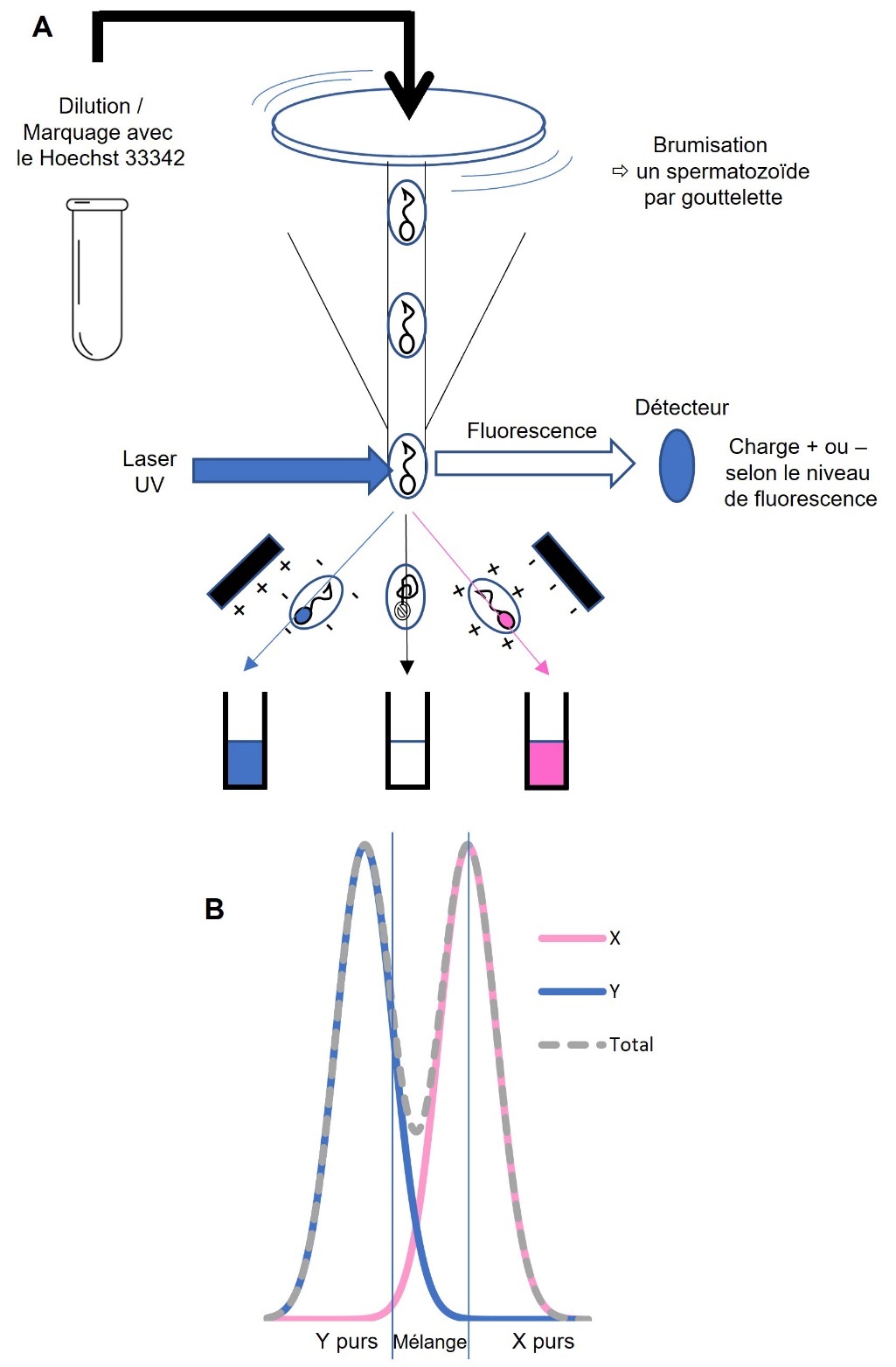
Inventée dans les années 1990, cette technique a largement profité des avancées des technologies de tri cellulaire, avec une augmentation du débit et une diminution du coût, et elle a pu être commercialisée depuis 2004. Une tête de trieur actuel permet d’obtenir environ 6 000 spermatozoïdes triés par seconde, un débit qui permet la production de quelques centaines de doses par machine et par jour. Les doses produites avec le protocole usuel contiennent moins de spermatozoïdes que les doses conventionnelles (environ 2 millions vs 8 millions), ce qui résulte également d’un optimum économique du process. Selon Sexing Technologies, qui reste discret sur la nature des améliorations, le procédé le plus récent, appelé sexedULTRA®, préserve mieux l’intégrité des spermatozoïdes et doit réduire la perte de fertilité. Récemment est apparue la possibilité technique d’augmenter le nombre de spermatozoïdes à 4 millions par paillette, ce qui doit également contribuer à réduire la perte de fertilité, mais à un coût plus élevé.
Les semences sexées usuelles garantissent 90 à 92 % de descendants du sexe demandé. L’essentiel des besoins concerne la production de doses femelles pour insémination dans les élevages commerciaux. Aujourd’hui, trois ateliers de sexage de la semence bovine fonctionnent en France. Propriétés de l’entreprise Sexing Technologies, ils sont installés dans trois centres d’insémination dans l’est (Roulans, 25), l’ouest (St Aubin du Cormier, 35) et plus récemment le nord de la France (Douai, 59). D’autres ateliers localisés à l’étranger contribuent également à produire des doses sexées utilisées en France.
Il arrive que la semence de certains taureaux ne puisse pas être sexée. Ce phénomène est généralement dû à la présence d’un réarrangement chromosomique dans le génome du taureau. La méiose conduit alors à une forte proportion de spermatozoïdes au génome déséquilibré, avec deux copies ou zéro copie du segment chromosomique concerné (Jourdain et al., 2023). Cette variation de quantité d’ADN perturbe la distribution de la fluorescence qui n’est plus représentative de la distribution des chromosomes X et Y. Une impossibilité de sexage est donc un signal d’appel d’un possible réarrangement chromosomique. Un tel réarrangement est une tare rare mais sévère, induisant une perte de fertilité mâle d’abord, puis une perte de fertilité femelle chez les filles qui l’ont reçu, et justifie la non-utilisation du taureau en insémination.
2. Diffusion de la semence sexée
2.1. En France
Un bilan complet d’utilisation de la semence sexée a été réalisé par Idele (Dominique, 2023). Nous renvoyons le lecteur à cette étude pour éviter les redondances et nous ne reprenons que quelques faits marquants. L’utilisation de la semence sexée a décollé à partir de 2010, après l’installation du premier laboratoire de sexage en France à Roulans (25). Depuis 2015, elle est relativement stable autour de 12 % des inséminations premières et 9 % des inséminations totales, et a présenté d’abord une légère décroissance suivie d’une croissance modérée depuis 2019 (figure 2).
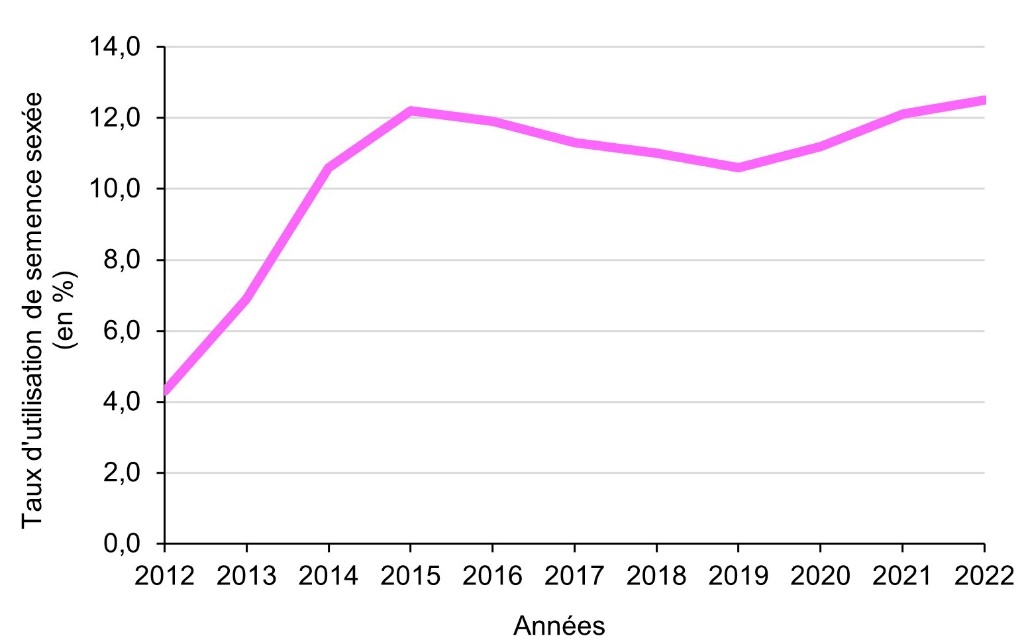
Ce bilan montre des différences très marquées. Tout d’abord, logiquement (comme expliqué en partie 4), la pratique du sexage est très limitée en races allaitantes (quelques %) alors qu’elle est beaucoup plus développée en races laitières. Elle est beaucoup plus importante sur les génisses que sur les vaches, du fait de leur fertilité plus élevée et d’une certaine sécurisation des conditions de naissance. Elle est plus développée sur la première insémination que sur les retours, 76 % des IA sexées étant des IA premières. Elle est très variable entre races : la race jersiaise présente le taux d’utilisation le plus élevé (60 % sur les génisses, 40 % sur les vaches), afin de limiter la naissance de veaux mâles dont la valorisation est quasi nulle ; les races brune et montbéliarde viennent ensuite, avec des niveaux élevés sur les génisses (45 % et 32 %) mais aussi sur les vaches (20 %) ; en races holstein et normande, le niveau d’utilisation est assez élevé en génisses (35 % et 23 %) mais faible en vaches (5 %) ; enfin, les races régionales ont des taux d’utilisation relativement bas. Globalement, les IA sexées représentent 9 % (583 000) des IA totales et 12,5 % des IA premières.
Le surcoût affiché par les entreprises d’insémination varie entre 18 et 25 € par dose. Ce montant assez élevé, associé à la perte de fertilité observée, constitue sans aucun doute un frein à l’utilisation et motive le ciblage des femelles les plus fertiles.
2.2. Situation internationale
Les premiers laboratoires ont été installés aux USA, au Royaume-Uni et en Suisse au début des années 2000. Depuis, leur nombre a augmenté et Sexing Technologies propose la même organisation dans chaque pays. Il dispose actuellement de 52 laboratoires de sexage répartis dans 20 pays, installés dans des centres de production de semence pour lesquels ils travaillent en prestation. Chaque laboratoire dispose généralement de plusieurs trieurs voire, pour les plus importants, de plusieurs dizaines de trieurs de cellules. La production mondiale de doses était estimée en 2018 à plus de 10 millions par an (González-Marín et al., 2018). Les résultats d’efficacité de sexage et de fertilité sont comparables dans tous les pays (Reese et al., 2021).
Le sexage de la semence permet également de répondre à des attentes sociétales de pays comme l’Inde où les veaux mâles ne peuvent pas être valorisés par des filières bouchères et restent toute leur vie une charge là où les vaches servent à la production laitière, apportant ainsi une source alimentaire et un complément de revenu important, dans un contexte de taille souvent très réduite des élevages.
3. Conséquences zootechniques de l’utilisation de semence sexée
L’intérêt majeur de la technique pour l’éleveur est évident et cette approche renouvelle considérablement la pratique de l’insémination et les stratégies de production de descendants. Elle permet de garantir le sexe du produit avec un haut niveau de fiabilité et donc de produire les génisses de renouvellement à partir de la partie du troupeau choisie par l’éleveur. En pratique, les allégations commerciales sont effectivement vérifiées : l’étude de la base de données nationale bovine hébergée au Centre de traitement de l’information génétique d’INRAE (CTIG, Jouy-en-Josas, https://ctig.jouy.hub.inrae.fr/) montre un taux de naissances femelles issues de semences sexées femelle de 91 %. Cette valeur a peu varié au cours du temps.
3.1. Conséquences sur la fertilité
Le procédé de sexage correspond à une manipulation lourde et assez longue qui a des conséquences sur la fertilité. Dans un premier temps, la semence est traitée au Hoechst 33342 qui a une certaine cytotoxicité. Même si seuls les spermatozoïdes vivants sont conservés après la phase de tri, il est probable que ce produit altère la survie ultérieure des spermatozoïdes triés. Par ailleurs, le traitement d’un éjaculat dure plusieurs heures, un délai que ne subit pas une semence conventionnelle. Le traitement implique des étapes de dilution, stress de température pendant le marquage, tri sous haute pression, exposition aux rayons laser UV, puis reconcentration. Enfin, pour des raisons économiques et pratiques, le nombre final de spermatozoïdes contenu dans une dose sexée est nettement inférieur à celui d’une dose conventionnelle, même en prenant en compte l’élimination des morts dans le processus de sexage. En 2009, Frijters et al. (2009) attribuaient près des deux tiers de la baisse de fertilité à la concentration plus faible et un tiers au procédé lui-même. Les évolutions récentes du procédé de sexage et la possibilité d’augmenter le nombre de spermatozoïdes par paillette doivent contribuer à limiter cette perte de fertilité.
L’analyse de la base nationale permet de mesurer l’impact du sexage sur la fertilité. Le taux de réussite moyen des inséminations artificielles conventionnelles varie d’environ 40 à 65 % selon la race et la parité. De nombreux facteurs affectent le taux de réussite et il convient de bien distinguer l’effet du sexage des autres effets. Le modèle utilisé en évaluation génétique intègre le statut de la semence (conventionnelle ou sexée) parmi de nombreux autres facteurs (troupeau, année, saison, parité, inséminateur, taureau, âge, intervalle mise bas insémination, valeur génétique de la femelle en fertilité…). La prise en compte de ces facteurs permet de limiter les biais liés à des confusions partielles entre effets et d’estimer l’effet propre du sexage de la semence. Dans les trois principales races laitières, la perte de fertilité est systématique et assez importante (–6 à –10 points de réussite à l’IA selon les races, les années, et la parité) comme le montre la figure 3. Les résultats se sont un peu améliorés dans le temps, surtout chez les vaches holstein et normandes, et la perte n’était plus que de six à huit points ces dernières années. De même, la différence de perte entre génisses et vaches, initialement forte, a pratiquement disparu aujourd’hui. Dans les races régionales, les effectifs sont plus réduits, mais la perte semble légèrement plus faible, comprise entre cinq et sept points.
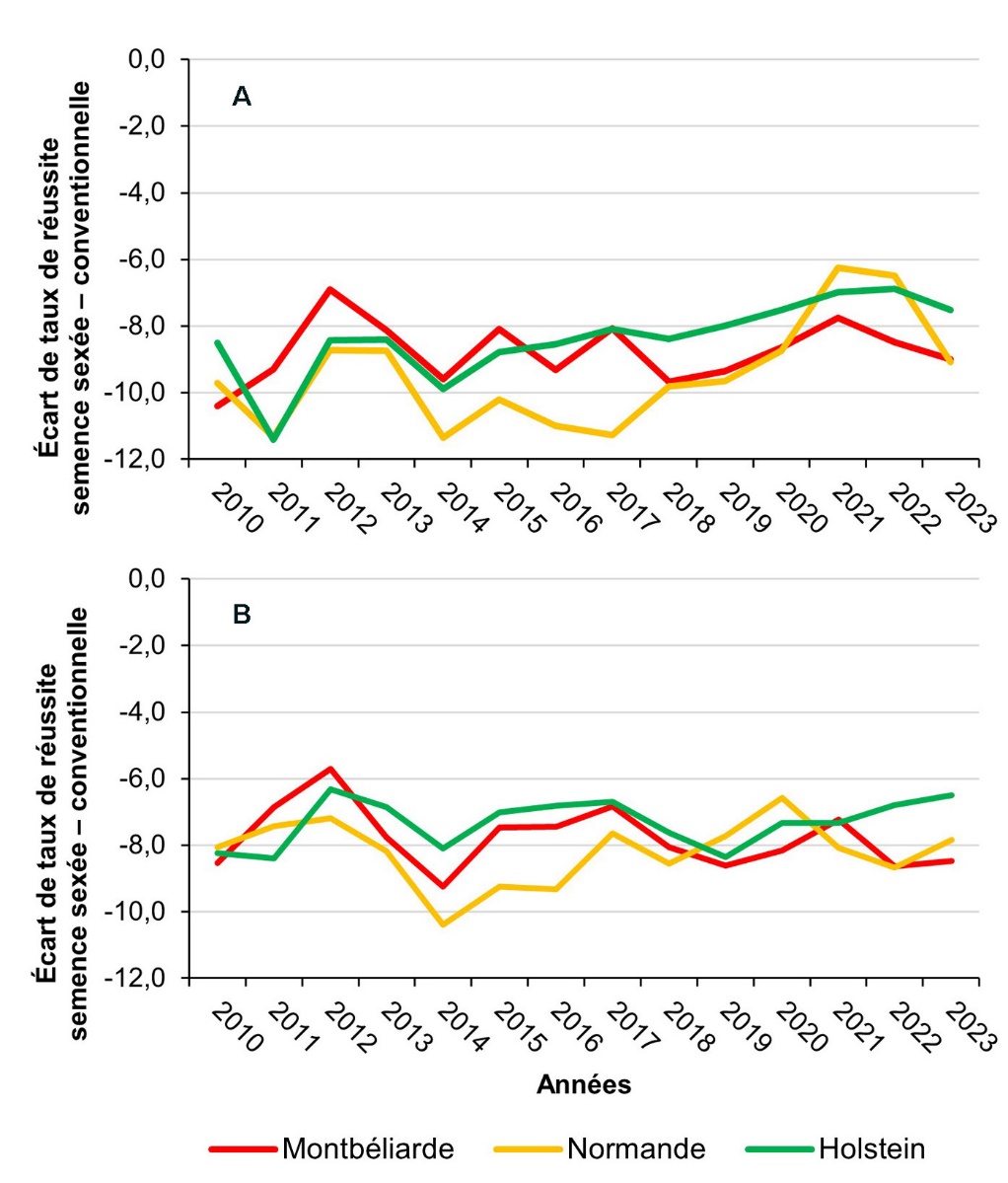
Cette moindre fertilité de la semence sexée est un effet bien connu, comme le montre la méta-analyse de Reese et al. (2021). En conséquence, compte tenu de son surcoût et d’un souhait de maintenir une bonne fertilité du troupeau, la semence sexée est utilisée préférentiellement dans les conditions de fertilité maximale. Elle est donc plutôt pratiquée à la première ou lors des deux premières inséminations, les IA ultérieures correspondant à des vaches nettement moins fertiles. En Holstein, elle est utilisée plutôt chez les génisses que chez les vaches en lactation, alors que cette stratégie est moins nette en Montbéliarde. Cette différence peut être attribuée à une meilleure fertilité dans cette race, mais aussi au marché de génisses amouillantes dynamique.
3.2. Conséquences sur les produits nés
L’impact sur les produits nés est moins connu et en général supposé nul ou négligeable. Il convient cependant de le vérifier, compte tenu de la large diffusion de la semence sexée. Des études ont été réalisées pour répondre à deux questions : i) le procédé de sexage respecte-t-il l’intégrité du génome du produit né ? En effet, le Hoechst 33342 se fixe sur l’ADN et pourrait donc induire des mutations de novo. ii) plus généralement, le procédé de sexage impacte-t-il les performances des produits nés ?
Le pouvoir mutagène du Hoechst 33342 fait débat. Pour Johnson & Schulman (1994), ce produit n’est pas un intercalant de l’ADN (au sens qu’il ne se fixe pas à l’intérieur de la double hélice) et sa fixation est réversible. De ce fait, il ne serait pas dangereux. Gardiner-Garden (1999), lui, indique qu’il peut déstabiliser les histones de l’ADN et affecter sa compaction dans les spermatozoïdes. Selon Parrilla et al. (2004), il n’a pas d’effet génotoxique sur la semence de verrat. Tirant profit des avancées des technologies de séquençage, des travaux de séquençage du génome ont été conduits en France en 2013-2014 dans le cadre du projet SexSeq financé par APIS-GENE pour répondre à la première question et rechercher les mutations de novo. Une mutation de novo est un variant observé avec certitude chez le produit et absent du génome de ses deux parents. Il existe un taux de mutation naturelle, estimé à environ 10-8 mutation par base d’ADN et par méiose chez les mammifères, soit environ 60-80 néomutations par produit né, et ces néomutations constituent la source originelle de la variabilité génétique. La question étudiée n’était pas de mesurer précisément le taux de néomutations supplémentaires liées au sexage (ce qui aurait demandé un dispositif potentiellement de très grande taille et très coûteux), mais simplement de savoir s’il était très fortement modifié, par exemple multiplié par 2, ou 10, voire plus. Compte tenu de cet objectif, le dispositif considéré était donc relativement petit et ne pouvait identifier que de très grosses différences. En effet, il impliquait deux familles, une de race holstein et l’autre de race montbéliarde. Chaque famille était constituée d’un taureau d’insémination, de deux vaches accouplées à ce taureau, et de deux filles par vache, l’une issue de semence conventionnelle et l’autre issue de semence sexée. Au total, le génome de quatre vaches issues de semence sexée a donc été comparé intrafamille au génome de quatre vaches issues de semence conventionnelle. Pour l’un des pères, la semence sexée a été produite après la semence conventionnelle, tandis qu’elle a été produite avant pour l’autre père, ce qui permet d’éviter une confusion entre les effets du type de semence et de l’âge du taureau. Le séquençage du génome des 14 animaux a été réalisé à une profondeur relativement élevée (25× minimum) pour éviter de confondre néomutation et erreur de séquençage. En moyenne, 80 à 120 mutations de novo ont été observées par individu indépendamment du type de semence, soit légèrement plus que ce qui était prédit par la bibliographie. Le nombre de néomutations a augmenté avec l’âge du taureau au moment de sa collecte, ce qui est un phénomène connu et vraisemblablement dû à l’accumulation des mitoses dans la spermatogenèse. Par contre, aucune différence significative n’a été observée entre les sœurs issues de semence normale et conventionnelle. Ce dispositif de taille réduite ne pouvait détecter que de grosses différences, et nous pouvons conclure que ce n’est pas le cas. Par contre, nous ne pouvons pas exclure des différences réduites, mais qui n’auraient pas de conséquences pratiques. En effet ces différences seraient inférieures à celles imputables à l’âge du taureau ou à un turnover plus rapide des générations (qui augmente le nombre de méioses et donc le nombre de mutations naturelles par an). Ces travaux, restés non publiés, ont permis de s’assurer que la large diffusion de la semence sexée ne s’accompagnait donc pas de risque spécifique de fardeau génétique additionnel pour le futur.
Des travaux récents (Fouéré et al., 2024) ont permis de répondre à la seconde question. Ils ont recherché un éventuel impact de la semence sexée sur les performances futures du veau né. Une large gamme de caractères a pu être étudiée à partir de la base de données zootechniques nationale : production laitière, mammites, stature en 1re lactation, fertilité en génisse et en 1re lactation. Pour éviter les risques de confusion, le modèle d’analyse a inclus les principaux effets de milieu considérés en évaluation génétique (dont un effet troupeau × année pour assurer une comparaison entre contemporains). L’étude a été limitée aux vaches génotypées et avec des premières lactations après 2018, ce qui a permis d’inclure leur valeur génomique dans le modèle. Dans ce modèle assez original, la valeur génomique n’est pas estimée car elle absorberait une grande partie de l’effet recherché. Elle est donc ajoutée en covariable dans le modèle : l’effet semence sexée mesure donc l’écart de performance par rapport à la valeur attendue, en limitant fortement le risque de confusion d’effet (par exemple, si la semence sexée est utilisée sur les meilleures vaches). La valeur génomique était celle estimée en 2018 et n’incluait donc pas les performances propres des animaux. Pour disposer d’effectifs génotypés suffisants, l’étude a été limitée aux races holstein et montbéliarde. Pour la plupart des caractères, les résultats ne montrent aucun effet (tableau 1). Le seul résultat significatif est un effet légèrement défavorable de la semence sexée sur la production laitière en race montbéliarde (–52 kg lait sur la lactation), non observé chez la Holstein. Le dispositif expérimental utilisé par Maicas et al. (2020) est plus restreint, mais il étudie des descendants de semence sexée ou conventionnelle issus d’un même éjaculat. Leurs conclusions sont similaires, pas de différence observée concernant la survie au cours de la première année de vie, les performances de production et de reproduction entre les descendants issus de semence sexée ou conventionnelle. S’ils existent, les effets de la semence sexée sont donc réduits et ont peu d’importance pratique.
Races | Holstein | Montbéliarde |
Taille1 | 0,00 (0,01) | 0,09 (0,06) |
Quantité de lait (kg) | –10,9 (9,4) | –51,8 (17,6)* |
Quantité de matière grasse (kg) | 0,21 (0,36) | –1,45 (0,67)* |
Quantité de matière protéique (kg) | –0,35 (0,29) | –1,92 (0,57)* |
Score de cellules somatiques | 0,00 (0,01) | 0,03 (0,02) |
Occurrences de mammites cliniques | 0,00 (0,003) | 0,00 (0,01) |
Taux de réussite à l’IA des génisses | 0,01 (0,004) | 0,01 (0,01) |
Taux de réussite à l’IA en L1 | 0,01 (0,01) | 0,04 (0,05) |
4. Intérêt de la semence sexée pour l’éleveur
L’intérêt majeur de la technique pour l’éleveur est évident en production laitière et cette approche renouvelle considérablement la pratique de l’insémination et les stratégies de production de descendants. Elle permet de garantir le sexe du produit avec un haut niveau de fiabilité et donc de produire les femelles de renouvellement sur une fraction choisie des vaches du troupeau, tandis que le reste du troupeau peut être conduit avec une autre politique de reproduction.
Elle présente cependant plusieurs inconvénients : i) un coût élevé, expliqué par la perte de semence induite, la lourdeur des investissements, le relativement faible débit, et le niveau élevé des tarifs imposé par Sexing Technologies en situation de monopole ; ii) une disponibilité limitée en nombre de doses et en diversité de taureaux, du fait à nouveau de la lourdeur du procédé et de son débit encore limité ; iii) enfin, la perte de fertilité décrite plus haut.
En races à viande, l’intérêt du sexage est plus limité. En race pure, les veaux mâles sont très bien valorisés. La bonne longévité limite le besoin de renouvellement et permet de cibler des filles de femelles confirmées. Le gain lié à une naissance plus facile des veaux femelles chez les génisses n’est pas suffisant pour compenser le surcoût de l’insémination et la baisse de fertilité de la semence sexée, dans un contexte général de faible pénétration de l’insémination dans ces populations. Notons également qu’en croisement sur support laitier, l’intérêt du sexage, qu’il soit femelle ou mâle, ne présente pas d’intérêt évident et le marché est inexistant.
Nous décrivons d’abord l’intérêt du sexage seul, puis en combinaison avec d’autres techniques.
4.1. Sexage seul
Le sexage seul (non combiné à d’autres techniques) permet le choix du sexe du veau issu d’un accouplement donné. Il augmente donc considérablement la probabilité d’obtenir une génisse d’une mère donnée. Par exemple, sous l’hypothèse d’un taux de mise bas de 50 et 44 % après une IA, d’un pourcentage de sexe femelle de 49 et 90 % avec semence normale ou sexée, et l’utilisation de semence sexée sur les deux premières inséminations seulement (la troisième étant en semence conventionnelle), la probabilité d’obtenir un veau de sexe femelle après trois IA maximum passe de 43 à 69 % (+62 %). Le renouvellement devient donc davantage choisi que subi.
La modification du sex-ratio induit deux autres conséquences qu’il est intéressant de mentionner. Les veaux femelles sont moins lourds et induisent des conditions de naissance plus faciles, en particulier chez les génisses. Ce point est d’autant plus notable que les inséminations avec semence sexée visent préférentiellement les génisses. De façon plus anecdotique, la durée de gestation des veaux femelles est inférieure d’un jour à celle des mâles, ce qui diminue d’autant l’intervalle entre vêlages.
En l’absence d’autres mesures, et même si l’utilisation de semence sexée se limite aux besoins de renouvellement, le troupeau produit un excès de veaux femelles (car les IA normales produisent aussi des femelles). Cela permet une sélection plus intense des femelles de renouvellement, mais sur la base des index sur ascendance uniquement, donc de façon relativement peu précise en l’absence de génomique. La procréation de femelles en excès offre plusieurs opportunités de valorisation. Ces femelles peuvent être vendues jeunes, ou élevées et vendues gestantes, mais le marché initialement favorable à cette activité a été rapidement saturé par un fort excès de génisses disponibles. Elles peuvent aussi être élevées pour la boucherie, mais l’intérêt est plus limité. Elles peuvent aussi, bien sûr, être conservées pour alimenter une croissance de l’effectif du troupeau, si c’est la politique recherchée. Par contre, il est généralement contre-productif d’accélérer artificiellement le renouvellement du troupeau pour intégrer cet excès de femelles, le coût d’une réforme prématurée d’une vache en cours de carrière est en effet très élevé. L’intérêt de l’utilisation de la semence sexée n’est donc marqué que si son utilisation s’accompagne d’autres changements. Pour conclure, malgré son surcoût, l’utilisation de la semence sexée sur une partie du troupeau est facilement rentabilisée par l’éleveur car, avec la quasi-garantie d’obtenir une femelle, elle permet une souplesse et une optimisation de la gestion du troupeau.
Le cas de la filière bio appelle une remarque. En effet, le sexage de la semence peut être considéré comme un processus non naturel. Par ailleurs, l’utilisation du Hoechst 33342 pose parfois question : cet intrant est-il compatible avec le cahier des charges de l’Agriculture biologique ? La question a émergé très tôt, dès les débuts de l’utilisation de la semence sexée. Elle a été tranchée au début des années 2010, tant en France qu’à l’étranger (en Suisse ou en Allemagne, par exemple). Aujourd’hui, cette technique est autorisée en bio mais la question est régulièrement revisitée.
4.2. Le sexage en combinaison avec le croisement et le génotypage en système laitier
L’utilisation de la semence sexée voit son intérêt fortement augmenter si l’excédent de femelles est maîtrisé d’une part, et si les choix de sélection pour assurer le renouvellement sont plus précis d’autre part.
Concernant le premier point, une fois les besoins de renouvellement assurés par l’emploi de la semence sexée, les vaches non retenues peuvent être accouplées avec des taureaux de race à viande pour produire des veaux croisés (mâles ou femelles) qui seront vendus à 15 jours ou élevés pour la viande. Cette politique est intéressante pour l’éleveur car la valorisation de ces veaux croisés est très bonne et le coût spécifique de reproduction est réduit. Le gain associé à ce coproduit couvre largement le surcoût des doses sexées. Cette pratique de croisement, avec des races variées, spécialisées ou culardes, s’est très fortement développée ces dernières années, en combinaison ou non avec la semence sexée, mais la synergie avec l’utilisation de la semence sexée est très forte. Ainsi, en 2022, le croisement terminal, en vue de la production de viande représentait 26 % des inséminations sur femelles laitières.
Au sein des systèmes laitiers avec une conduite en vêlages groupés, tels les systèmes herbagers, la réussite de la courte saison de reproduction (12 à 14 semaines, à dates fixes) est un challenge lourd de conséquences en matière de longévité des vaches, les vaches vides étant réformées en fin de lactation. L’utilisation de la semence sexée en début de saison de reproduction, permet d’assurer tôt le renouvellement et ensuite, d’utiliser des taureaux de race à viande en monte libre, en vue surtout de garantir la gestation de la majorité des vaches, tout en produisant des animaux croisés (de Vries et al., 2008 ; Holden & Butler, 2018 ; Berry, 2021). Ces animaux croisés trouvent aussi leur pertinence dans la valorisation de surfaces en prairies éloignées de la salle de traite, et/ou de moindre qualité floristique ou agronomique. Cette stratégie est bien illustrée par l’expérimentation conduite au domaine expérimental INRAE du Pin (Delaby et al., 2024). Les vaches sont conduites en système herbager avec une saison de reproduction courte pour obtenir des vêlages très groupés. Après cinq semaines d'inséminations en semence sexée de race pure, les vaches sont ensuite inséminées ou saillies en croisement Angus. Les veaux nés de ce croisement sont élevés avec les génisses laitières jusqu’à cinq mois, puis sont conduits à l’herbe, finis à l’étable avec un très bon ensilage d'herbe et abattus à 22 mois.
Concernant le second point, le choix des vaches dédiées au renouvellement en race pure et de l’orientation des autres vers le croisement est considérablement amélioré par les prédictions génomiques. En effet, les évaluations génomiques sont très précises pour tous les caractères, même peu héritables, beaucoup plus précises qu’un simple index sur ascendance. Alors que la sélection bovine sur la voie mère-fille a été longtemps considérée comme très peu efficace par manque à la fois d’intensité de sélection et de précision, l’évaluation génomique après génotypage apporte la précision tandis que la semence sexée permet d’augmenter l’intensité de sélection. La sélection intratroupeau peut maintenant contribuer plus efficacement à la création de progrès génétique. Ainsi, avec une pression de sélection d’une vache sur deux pour procréer le renouvellement, une précision R des index génomiques d’au moins 0,7 sur l’objectif de sélection, les filles disposent d’une supériorité de l’ordre de 0,2 à 0,3 écart-type génétique, qu’elles exprimeront lorsqu’elles entreront en carrière productive. Et comme toujours en génétique, ce gain est acquis et cumulatif. En supposant un intervalle de génération de cinq ans, cette pratique permet d’augmenter le progrès génétique de 20 % environ, par rapport à une situation sans semence sexée ni génotypage intratroupeau, dans laquelle le progrès génétique est quasi exclusivement apporté par les taureaux.
Par ailleurs, les index génomiques étant disponibles et précis (avec la même précision pour les femelles que pour les taureaux) pour un grand nombre de caractères, le choix des mères du renouvellement peut être réalisé sur la base d’un objectif spécifique défini par l’éleveur en fonction de sa situation propre (par exemple, la composition du lait, la production, la morphologie, la résistance à certaines maladies…). Par exemple, dans des élevages avec une forte prévalence de paratuberculose, une sélection spécifique sur l’index de résistance à cette maladie contribuera à assainir le troupeau, et ce d’autant plus rapidement que la contamination potentielle du produit est très précoce dans le premier mois de vie. On peut aussi citer la filière bio dont les objectifs de sélection peuvent s’écarter des objectifs généraux. L’utilisation conjointe de sélection génomique et de semence sexée donne beaucoup plus de latitude pour définir un objectif particulier.
Alors que le croisement coûte peu et qu’il est rentabilisé rapidement (dès la naissance du produit), l’utilisation de semence sexée et le génotypage sont des investissements à plus long terme. Si le coût de l’utilisation de la semence sexée peut être financé par un nombre accru de veaux croisés, le gain apporté par le génotypage et donc par un progrès génétique plus élevé, est plus tardif : en effet, les premiers retours sur investissement ne sont attendus que lorsque les produits sont en production, soit environ trois à cinq ans plus tard. Selon Bérodier et al. (2019) l’investissement dans le génotypage est complètement rentabilisé en huit ans sur la base du progrès génétique supplémentaire acquis. Mais ces auteurs supposaient l’élevage de nombreuses génisses, ce qui n’est pas optimal. Compte tenu de la baisse du prix du génotypage et de la valeur de ses autres apports (gestion des anomalies, de la consanguinité, des accouplements…), de l’optimisation du renouvellement pour ne procréer et élever que le nombre nécessaire de génisses, et de situations spécifiques (par exemple, la réduction de la prévalence de maladies, comme mentionné plus haut), le retour total sur investissement est généralement nettement plus rapide.
Dans l’absolu, on pourrait imaginer deux stratégies de sélection intratroupeau, qui gèrent différemment le compromis entre court et long termes. En effet, à court terme, il est intéressant de maximiser le croisement et de ne produire que le renouvellement nécessaire, ce qui limite forcément la pression de sélection disponible plus tard et donc le progrès génétique à terme. Au contraire, la procréation d’un plus grand nombre de génisses de renouvellement permet un tri plus sévère après génotypage de celles qui sont conservées et un progrès génétique plus fort, mais au prix d’une moindre proportion de croisement et d’un excès de génisses pures. D’un point de vue économique, c’est donc la première solution qui doit être privilégiée car elle est rentable plus vite en maximisant le croisement, en limitant les frais d’élevage de génisses, et du fait d’un effectif à génotyper plus faible. D’un point de vue éthique et environnemental, limiter le renouvellement et favoriser des carrières plus longues est également hautement souhaitable.
En conclusion, c’est donc en combinant l’utilisation de la semence sexée avec le croisement et le génotypage que l’on maximise son intérêt. À l’avenir, on peut prédire un développement très fort de l’utilisation combinée de ces techniques, surtout si le handicap de moindre fertilité de la semence sexée est réduit ou résolu et/ou si les surcoûts de la semence sexée diminuent.
5. Conséquences de l’utilisation de la semence sexée sur la filière bovine
5.1. Production de génisses
Comme indiqué précédemment, l’utilisation de semence sexée a fortement augmenté le nombre de génisses laitières nées et disponibles sur le marché. Dans un premier temps, l’évolution a été bénéfique car il y avait un déficit structurel de génisses à vendre sur le marché français et les possibilités d’export étaient importantes. Ainsi, dans le cas particulier de la race jersiaise, la disponibilité de génisses a permis l’augmentation spectaculaire des effectifs qui ont été multipliés par trois en 10 ans. Mais rapidement, et dans toutes les races, le marché est arrivé à saturation, associé à un excédent structurel de génisses et une forte pression sur leur prix. Il convient donc de limiter la procréation de génisses aux besoins anticipés à la fois intra-élevage et pour la filière. Il n’en demeure pas moins que la semence sexée est un outil qui donne beaucoup de flexibilité pour accroître rapidement le nombre de femelles si le besoin s’en fait sentir.
5.2. Production de veaux croisés et impact sur la filière viande
Dans un second temps, le croisement « viande » s’est énormément développé. Plus le taux d’IA sexées est élevé, plus le pourcentage d’IA de croisement peut augmenter, potentiellement jusqu’à un maximum de 50 à 65 % selon les besoins de renouvellement. En 2022, 26 % des inséminations totales et 20 % des inséminations premières en races laitières ont été réalisées en croisement avec des races à viande, le maximum étant observé en race montbéliarde. Cette pénétration du croisement « viande » a plus que doublé en 10 ans, tirée en particulier par un quasi-triplement en race holstein. Cette pratique de croisement est favorable pour l’efficience technique et la rentabilité de l’élevage laitier avec un coproduit à bonne valeur ajoutée, dans un contexte où les veaux de race pure sont au contraire généralement mal valorisés, notamment certaines années ou à certaines saisons. Par contre, cela pourrait constituer une concurrence importante pour la filière viande, surtout si ces animaux sont à l’avenir élevés comme jeunes bovins, augmentant alors substantiellement le tonnage de viande produite à partir de la filière laitière. Le facteur « viande issue du lait » est donc à intégrer dans les scénarios d’avenir des filières bovines laitière et allaitante. Il n’aura pas le même impact selon les hypothèses de consommation de viande en volume et en nature (haché ou piècé) et d’effectifs nationaux des troupeaux laitiers et allaitants.
5.3. Impact sur la génétique
Les impacts sur la filière génétique sont assez paradoxaux. Les gains en termes de progrès génétiques bénéficient aux éleveurs, en particulier dans le scénario combinant sexage, croisement et génotypage qui augmente substantiellement le gain génétique et permet une certaine diversification des objectifs. À l’inverse, notons aussi que la semence sexée n’est pas disponible pour tous les taureaux mis en marché, ce qui réduit la diversité de l’offre.
Le bilan est plus mitigé pour les entreprises de sélection. La vente de semence sexée femelle assure une protection partielle et temporaire de la génétique des entreprises vis-à-vis de leurs concurrents. En effet, les entreprises de sélection ne commercialisent pas de semence sexée mâle, à la fois parce que les éleveurs n’en veulent généralement pas et parce qu’elles veulent éviter la production de mâles qui viendraient concurrencer leur activité. Par contre, elles l’utilisent en interne pour procréer leurs propres taureaux, particulièrement en fécondation in vitro. Mais cette utilisation est très limitée en volume.
Mais avec le sexage combiné au croisement, un point crucial est la diminution du nombre d’inséminations en race pure. Or, c’est le nombre d’inséminations en race pure qui assure l’équilibre économique des programmes de sélection. Les inséminations sexées sont vendues plus cher mais coûtent aussi davantage, et la plus-value bénéficie surtout à Sexing Technologies plutôt qu’aux entreprises de sélection. Par ailleurs, Sexing Technologies limite le nombre de laboratoires installés et toutes les entreprises de sélection ne peuvent pas en héberger, ce qui les oblige à réaliser le sexage dans des laboratoires distants et sur des sites potentiellement concurrents. Si l’entreprise de sélection souhaite éviter d’acheter des doses de taureaux à viande pour le croisement, elle doit les produire elle-même, c’est-à-dire conduire des programmes de sélection supplémentaires et réaliser les investissements correspondants. Enfin, il convient de noter que le développement de la semence sexée finance confortablement une entreprise en situation de monopole qui, en se diversifiant, est devenue un acteur de premier plan en sélection et vente de génétique, constituant une menace nouvelle et majeure pour les entreprises de sélection historiques.
Conclusion
Toutes les entreprises de sélection bovine, tout particulièrement en filière laitière, proposent de la semence sexée femelle à leur catalogue. Depuis 2010, la proportion d’inséminations avec semence sexée augmente progressivement. La croissance est assez lente du fait du surcoût de la semence sexée, de la baisse de fertilité qui, au moins en race holstein, en oriente l’usage principalement sur les génisses, et sur le partage inégal de la valeur ajoutée du procédé au détriment des entreprises de sélection. L’augmentation observée devrait continuer et s’amplifier car le niveau actuel de parts de marché des semences sexées (moins de 10 % du total des inséminations) est très en dessous de l’optimum technique. Le développement fort et régulier du croisement viande et du génotypage devrait constituer un moteur puissant car c’est en combinant ces trois techniques que l’éleveur tire le maximum de bénéfice. Le développement du croisement est une évolution majeure pour la valorisation des produits viande issus du secteur laitier, mais constitue une menace potentielle pour le secteur allaitant, en fonction des scénarios d’évolution envisagés.
Contribution des auteurs
DB a conçu l’article et a écrit sa version initiale ; SD a réalisé le bilan d’utilisation de la semence sexée ; MB, SF et LD ont modélisé l’impact du sexage, du croisement et du génotypage ; CF a estimé l’effet de la semence sexée sur les performances des filles ; MB a estimé le taux de mutation de novo associé à la semence sexée ; AB a estimé la perte de fertilité associée à l’utilisation de semence sexée ; tous les auteurs ont contribué à la rédaction et approuvent le manuscrit.
Remerciements
APIS-GENE a financé le projet SEXSEQ sur la recherche de néomutations associées à l’utilisation de semence sexée. Les travaux sur les effets de programmation associés à la semence sexée ont été réalisés dans le cadre du projet POLYPHEME financé par APIS-GENE et l’Agence nationale de la recherche (ANR-21-CE20-0021). Idele remercie le CASDAR pour le financement des bilans de l’utilisation des semences sexées.
Références
- Bérodier, M., Brochard, M., Boichard, D., Dezetter, C., Bareille, N., & Ducrocq, V. (2019). Use of sexed semen and female genotyping impacts genetic and economic outcomes of Montbéliarde dairy herds depending on the farming system considered. Journal of Dairy Science, 102(11), 10073-10087. doi:10.3168/jds.2018-16041
- Berry, D. P. (2021). Invited review: Beef-on-dairy. The generation of crossbred beef x dairy cattle. Journal of Dairy Science, 104(4), 3789-3819. doi:10.3168/jds.2020-19519
- Boichard, D. (2020). Nouvelles modalités de diffusion de la génétique. In E. Verrier, D. Milan, & C. Rogel-Gaillard (Coords.), Génétique des animaux d'élevage : Diversité et adaptation dans un monde changeant (pp. 193-203). Versailles : Éditions Quae. https://www.quae.com/produit/1635/9782759231003/genetique-des-animaux-d-elevage
- Delaby, L., Bachelet, A., Gaborit, M., Launay, F. (2024). Tripl'Scotch: Beef from dairy cows managed within a compact spring calving grass-based system in Western France. In E. Fitzpatrick (Ed.), Dairy calf-to-beef Conference proceedings (pp. 88-89). Wexford: Teagasc . https://www.teagasc.ie/publications/2024/dairy-calf-to-beef-conference.php
- De Vries, A., Overton, M., Fetrow, J., Leslie, K., Eicker, S., & Rogers, G. (2008). Exploring the impact of sexed semen on the structure of the dairy industry. Journal of Dairy Science, 91(2), 847-856. doi:10.3168/jds.2007-0536
- Dominique, S. (2023). Bilan des inséminations animales bovines en semence sexée, campagne 2022. Consulté le 2 septembre 2024 sur Idele https://idele.fr/detail-article/bilan-des-inseminations-animales-bovines-en-semence-sexee-campagne-2022
- Druart, X. & Ribeiro Bento Dos Santos, C. (2004). Le sexage des spermatozoïdes : état des lieux et perspectives. Rencontres Recherches Ruminants, 11, 369-372. https://www.journees3r.fr/spip.php?article574
- Foote, R. H. (2002). The history of artificial insemination. Selected notes and notables. Journal of Animal Science, 80(suppl 2), 1-10. doi:10.2527/animalsci2002.80E-Suppl_21a
- Fouéré, C., Sanchez, M. P., Boussaha, M., Fritz, S., Vinet, A., Kiefer, H., Boichard, D., & Hozé, C. (2024). A large population study to assess the magnitude of prenatal programming in dairy cattle. Journal of Dairy Science, 107(8), 5913-5923. doi:10.3168/jds.2023-24051
- Frijters, A. C. J., Mullaart, E., Roelofs, R. M. G., van Hoorne, R. P., Moreno, J. F., Moreno, O., Merton, J. S. (2009). What affects fertility of sexed bull semen more, low sperm dosage or the sorting process? Theriogenology, 71(1), 64-67. doi:10.1016/j.theriogenology.2008.09.025
- Gardiner-Garden, M. (1999). Techniques for sorting X and Y spermatozoa may adversely affect histone-associated regions in human spermatozoa. Human Reproduction, 14(5), 1403-1404. doi:10.1093/humrep/14.5.1403
- González-Marín C., Góngora, C. E., Gilligan, T. B., Evans, K. M., Moreno, J. F., & Vishwanath, R. (2018). In vitro sperm quality and DNA integrity of SexedULTRATM sex-sorted sperm compared to non-sorted bovine sperm. Theriogenology, 114, 40-45. doi:10.1016/j.theriogenology.2018.03.025
- Holden, S. A, & Butler, S. T. (2018). Review: Applications and benefits of sexed semen in dairy and beef herds. Animal, 12(Suppl. 1),97-103. doi:10.1017/S1751731118000721
- Johnson, L. A. (1992). Methods to preselect the sex offspring (US5135759A). United States Patent. https://patentimages.storage.googleapis.com/2c/24/5d/47286fc1ea83ff/US5135759.pdf
- Johnson, L. A., Flook, J. P., & Look, M. V. (1987). Flow-cytometry of X and Y chromosome-bearing sperm for DNA using an improved preparation method and staining with Hoechst-33342. Gamete Research, 17(3), 203-12. doi:10.1002/mrd.1120170303
- Johnson, L. A., & Schulman, L. D. (1994). The safety of sperm selection by flow cytometry. Human Reproduction, 9(5), 758-759. doi:10.1093/oxfordjournals.humrep.a138591
- Johnson, L. A., & Siedel, G. E. Jr. (1999). Proceedings of the symposium current status of sexing mammalian sperm. Theriogenology, 52(8), 1267-1481. https://www.sciencedirect.com/journal/theriogenology/vol/52/issue/8
- Jourdain, J., Barasc, H., Faraut, T., Calgaro, A., Bonnet, N., Marcuzzo, C., … Capitan, A. (2023). Large-scale detection and characterization of interchromosomal rearrangements in normozoospermic bulls using massive genotype and phenotype datasets. Genome Research, 33, 957-971. doi:10.1101/gr.277787.123
- Maicas, C., Hutchinson, I. A., Cromie, A. R., Lonergan, P., & Butler, S.T. (2020). Characteristics of offspring derived from conventional and X-sorted bovine sperm. Journal of Dairy Science, 103(8), 7509-7520. doi:10.3168/jds.2020-18178.
- Parrilla, I., Vázquez, J. M., Cuello, C., Gil, M. A., Roca J., Di Berardino, D., & Martínez, E. A. (2004). Hoechst 33342 stain and UV laser exposure do not induce genotoxic effects in flow-sorted boar spermatozoa. Reproduction, 128(5), 615-621. doi:10.1530/rep.1.00288
- Reese, S., Camara Pirez, M., Steele, H., & Kölle, S. (2021). The reproductive success of bovine sperm after sex‑sorting: a meta‑analysis. Scientific Reports, 11, 17366. doi:10.1038/s41598-021-96834-2
- Umehara, T., Tsujita, N., Zhu, Z., Ikedo, M., & Shimada, M. (2020). A simple sperm-sexing method that activates TLR7/8 on X sperm for the efficient production of sexed mouse or cattle embryos. Nature Protocols, 15, 2645-2667. doi:10.1038/s41596-020-0348-y
Résumé
Le sexage de la semence de taureau (également validé chez diverses espèces de mammifères) est un procédé permettant de séparer les spermatozoïdes porteurs du chromosome X de ceux porteurs du Y. Le procédé actuel diffusé depuis 20 ans par l’entreprise Sexing Technologies repose sur le tri cellulaire, basé sur le contenu en ADN plus élevé pour les porteurs du X que du Y. La précision du sexage est d’environ 91 %. La part de marché de la semence sexée (femelle) atteint actuellement 12,5 % des premières inséminations. Son utilisation est très majoritairement en races laitières et plutôt sur génisses. Elle est la plus élevée en races jersiaise, montbéliarde et brune. Par rapport à la semence conventionnelle, elle s’accompagne d’une baisse de fertilité de l’ordre de 6 à 8 points de réussite, baisse qui tend à se réduire avec le temps. Aucun autre effet défavorable n’est observé. Le produit né ne présente pas d’augmentation du taux de néomutation dans son génome, ni de perte de performance significative sur aucun des caractères analysés. L’intérêt principal de la semence sexée se concrétise dans l’utilisation conjointe avec deux autres outils : tandis que la semence sexée est réservée à la part du troupeau assurant le renouvellement, dont le choix est bien plus précis avec génotypage et évaluation génomique, le croisement avec un taureau de race à viande assure une meilleure valorisation des produits non conservés pour le renouvellement. Ces trois techniques devraient donc continuer à se développer, surtout si la perte de fertilité liée au sexage est limitée.
Pièces jointes
Pas de document complémentaire pour cet articleStatistiques de l'article
 Vues: 10065
Vues: 10065
Téléchargements
 PDF: 716
PDF: 716
 XML: 209
XML: 209
Articles les plus lus par le même auteur ou la même autrice
- Audrey MICHAUD, Sylvain PLANTUREUX, René BAUMONT, Luc DELABY, Les prairies, une richesse et un support d’innovation pour des élevages de ruminants plus durables et acceptables , INRAE Productions Animales: Vol. 33 No 3 (2020)
- Nathan BALANDRAUD, Claire MOSNIER, Luc DELABY, François DUBIEF, Jean-Philippe GORON, Bruno MARTIN, Dominique POMIES, Anaël CASSARD, Holstein ou Montbéliarde : des différences phénotypiques aux conséquences économiques à l’échelle de l’exploitation , INRAE Productions Animales: Vol. 31 No 4 (2018)
- Charlotte DEZETTER, Didier BOICHARD, Nathalie BAREILLE, Bénédicte GRIMARD, Pascale LE MEZEC, Vincent DUCROCQ, Le croisement entre races bovines laitières : intérêts et limites pour des ateliers en race pure Prim’Holstein ? , INRAE Productions Animales: Vol. 32 No 3 (2019)
- Aurélie MADRID, Renée DE CRÉMOUX, Luc DELABY, Hélène LARROQUE, Sandra NOVAK, Aurélie VINET, L’élevage de ruminants s’adaptera-t-il au changement climatique ? Impacts et leviers d’adaptation , INRAE Productions Animales: Vol. 38 No 2 (2025)
- Olivier GODINOT, Sylvain FORAY, Sophie LEMOSQUET, Luc DELABY, Nadège ÉDOUARD, De l’animal au territoire, regards sur l’efficience de l’azote dans les systèmes bovins laitiers , INRAE Productions Animales: Vol. 35 No 1 (2022)
- Marie-Pierre SANCHEZ, Valérie WOLF, Cécile LAITHIER, Mohammed EL JABRI, Éric BEUVIER, Odile ROLET-RÉPÉCAUD, Nicolas GAUDILLIÈRE, Stéphanie MINÉRY, Yuliaxis RAMAYO-CALDAS, Thierry TRIBOUT, Alexis MICHENET, Mekki BOUSSAHA, Sébastien TAUSSAT, Sébastien FRITZ, Agnès DELACROIX-BUCHET, Philippe GROSPERRIN, Mickaël BROCHARD, Didier BOICHARD, Analyse génétique de la « fromageabilité » du lait de vache prédite par spectrométrie dans le moyen infrarouge en race Montbéliarde , INRAE Productions Animales: Vol. 32 No 3 (2019)
- Hervé GUYOMARD, Zohra BOUAMRA-MECHEMACHE, Vincent CHATELLIER, Luc DELABY, Cécile DÉTANG-DÉSSENDRE, Jean-Louis PEYRAUD, Vincent RÉQUILLART, Pourquoi et comment réguler la production et la consommation de produits animaux ? Le cas de l'Union européenne , INRAE Productions Animales: Vol. 34 No 3 (2021)
- Pascal RAINARD, Gilles FOUCRAS, Didier BOICHARD, Rachel RUPP, Faibles concentrations cellulaires du lait et sensibilité aux mammites des ruminants laitiers , INRAE Productions Animales: Vol. 31 No 4 (2018)
- Laurianne CANARIO, Nicolas BÉDÈRE, Marc VANDEPUTTE, Didier BOICHARD, Jérôme RAOUL, Catherine LARZUL, Quelles génétiques pour les systèmes d’élevages certifiés en agriculture biologique ? , INRAE Productions Animales: Vol. 37 No 2 (2024): Numéro spécial : L’élevage biologique : conditions et potentiel de développement
- Luc DELABY, Brendan HORAN, Améliorer l'efficacité des systèmes laitiers herbagers en milieux tempérés , INRAE Productions Animales: Vol. 34 No 3 (2021)