

Réduction des émissions de méthane entérique chez les ruminants : enjeux, solutions et perspectives à l’échelle de l’animal et des systèmes d’élevage (Full text available in English)
Le méthane est un gaz à fort pouvoir réchauffant sur le climat. Les connaissances scientifiques et les technologies évoluent rapidement pour proposer de nouvelles stratégies pour réduire les émissions de méthane entérique par les ruminants. Les conséquences des changements de pratiques et d’objectifs de sélection génétique doivent être évaluées de manière holistique à l’échelle de l’animal, des systèmes d’élevage, et des territoires dans lesquels ils s’insèrent.
Introduction
Les enjeux climatiques s’accentuent d’années en années avec une combinaison de phénomènes aigus et chroniques (pics et vagues de chaleur, déficits ou fortes précipitations) dont la fréquence et la sévérité s’amplifient. Le niveau de réchauffement planétaire est projeté par les études du Groupe d’experts intergouvernemental sur l’évolution du climat (GIEC) à +2 °C à l’horizon 2050 et +3 à +4 °C en 2100 comparativement à l’ère préindustrielle (IPCC, 2023). Malgré l’existence de traités internationaux sur l'atténuation du changement climatique, les émissions de gaz à effet de serre (GES) liées aux activités humaines (anthropiques) continuent d’augmenter, bien que le rythme au cours de la dernière décennie soit deux fois moins rapide que lors de la décennie précédente (IPCC, 2023). L’élevage représente 12 % des émissions totales de GES d’origine anthropique (c’est-à-dire liée aux activités humaines). Les émissions de méthane entérique, un gaz issu de la digestion des ressources alimentaires par les ruminants, représentent un peu moins de la moitié des GES associés à l’élevage (FAO, 2023). Leur réduction est un objectif prioritaire des politiques climatiques du fait du fort pouvoir réchauffant du méthane et de sa courte demi-vie avant de se transformer en dioxyde de carbone (encadré 1). L’engagement lors de la COP26 est de réduire les émissions de méthane au niveau mondial de –30 % en 2030 par rapport à 2020, et de –50 % à horizon 2050.
Les principales sources des émissions de méthane (CH4) liées aux activités humaines sont l’élevage et la riziculture (méthane biogénique) ainsi que l’extraction, la distribution des combustibles fossiles et les sites d’enfouissement (méthane fossile).
Le potentiel de réchauffement global (PRG) à 100 ans des gaz à effet de serre (GES) se calcule par la formule PRG100 = CO2 + 27,8 CH4 biogénique (ou 29,8 CH4 fossile) + 273 N2O (IPCC, 2023). Le méthane biogénique équivaut ainsi à 28 fois le dioxyde de carbone (CO2), une contribution bien moindre que celle du protoxyde d’azote (N2O). Mais le méthane a une demi-vie de 10 à 12 ans, ce qui en fait un objectif prioritaire des politiques visant à obtenir une atténuation rapide du réchauffement planétaire. Autrement dit, l’effet réchauffant du méthane est très fort à court terme, mais il ne s’accumule pas dans l’atmosphère et des émissions décroissantes conduisent à un effet refroidissant.
Le PRG100 est actuellement utilisé par tous les calculs nationaux évaluant les émissions de GES. De nouvelles métriques comme le PRG* sont proposées pour mieux rendre compte de la physique du méthane et de sa courte durée de vie (Poux et al., 2025). Le PRG* reflète l’équivalence entre un niveau d’émission de méthane et les émissions cumulatives de CO2 pour les effets sur le climat, en considérant l’impact de la dynamique des émissions pendant 20 ans sur la température moyenne du globe. En Europe où les émissions de méthane sont en baisse, la prise en compte du PRG* conduirait à une vision plus équilibrée entre CH4 et N2O dans la définition d’une trajectoire multigaz issus du secteur agricole pour atteindre l’ambition « zéro émissions nettes » de CO2 (émissions moins les puits de carbone).
L’Europe est l’une des seules régions du globe où les émissions de méthane diminuent depuis 1990 (EEA, 2024). Les cheptels de ruminants y sont en diminution, soit du fait d’une politique de réduction volontariste pour des raisons économiques ou environnementales (réduction des pollutions azotées), soit consécutivement à l’augmentation de la productivité des vaches laitières plus rapide que l’évolution de la demande en lait, et du moindre renouvellement des générations d’éleveurs (CGAAER, 2024). Il est probable que cette baisse continue au cours des 15 prochaines années. Toutefois, cette réduction des cheptels ne peut être la seule voie de réduction des émissions de méthane, car elle fait l’impasse sur les aménités positives associées aux élevages de ruminants, telles que la fourniture de produits de qualité appréciés des consommateurs, le rôle positif des prairies et des parcours dans la séquestration du carbone, et le bouclage des cycles de nutriments. Une divergence entre production et consommation de produits animaux peut également conduire à déporter les sources d’émissions de méthane dans d’autres pays, sans bénéfice au niveau planétaire. De ce fait, le déploiement de pratiques ou de technologies visant à la réduction des émissions de méthane entérique par les ruminants est attendu ou annoncé par les différents pays européens (CGAAER, 2024), avec l’objectif de rendre les baisses d’émissions moins dépendantes de la baisse des cheptels et d’une réduction de la consommation de leurs produits. De nombreux leviers sont proposés, basés notamment sur les compléments alimentaires, la composition des rations, la conduite des troupeaux ou la sélection génétique. Cependant, le changement climatique modifie la disponibilité et la qualité de certaines ressources végétales et questionne l’adaptation de l’animal à l’environnement, induisant des réponses variables à ces leviers. De plus, les transitions en cours des modèles agricoles et alimentaires génèrent des opportunités mais aussi des contraintes parfois contradictoires pour les systèmes d’élevage et les territoires dans lesquels ils s’insèrent.
Plusieurs défis sont ainsi posés à la recherche agronomique et au développement agricole pour réduire les émissions de méthane entérique associées aux systèmes d’élevage de ruminants. Un premier défi est d’identifier des stratégies combinant plusieurs leviers déjà existants pour réduire les émissions de méthane entérique, mais tout en préservant les performances de production de l’animal (quantité et qualité des produits), sa santé et son bien-être. Un deuxième défi est d’identifier de nouveaux leviers de réduction des émissions de méthane entérique dans un contexte de dépendance accrue des systèmes agricoles aux conditions locales, elles-mêmes challengées par le changement climatique. Un troisième défi est d’identifier des solutions gagnantes-gagnantes sur l’ensemble des aménités des systèmes d’élevage de ruminants tout en considérant l’impact sur les émissions d’autres GES.
L’objectif de cet article de synthèse est de faire un état des lieux des travaux sur la réduction des émissions de méthane des ruminants menés au cours de la dernière décennie par INRAE et ses partenaires de la recherche et du développement. Le méthane entérique est produit par la dégradation des aliments (fourrages et concentrés) en acides gras volatils (AGV) par certains des microorganismes (archées méthanogènes) présents dans le rumen qui utilisent l’hydrogène pour leur métabolisme et leur croissance (figure 1).
Au sein du rumen, les voies impliquées dans la consommation de l’hydrogène (H2) sont représentées par des flèches entre compartiments, avec une intensité minimale pour la biomasse microbienne, moyenne pour la production de propionate (AGV : acides gras volatils) et maximale pour la production de méthane (CH4). Des enjeux scientifiques, adossés à une meilleure connaissance du fonctionnement du rumen et de l’animal hôte, sont indiqués sous forme de cartouches positionnées respectivement à droite et à gauche de la figure.
Figure 1. Composantes des émissions de méthane entérique et objectifs de recherche.
Évaluer l’efficacité de modifications alimentaires, génétiques ou de conduites plus globales des animaux nécessite de disposer d’outils performants pour mesurer les émissions de méthane entérique par l’animal, et de prédicteurs pour les évaluer dans différents contextes (partie 1). Identifier de nouvelles cibles d’actions implique de caractériser et comprendre la diversité de la biomasse microbienne, afin de moduler ses fonctions à des périodes clés de la vie de l’animal (partie 2). Élaborer des stratégies de réduction des émissions de méthane entérique nécessite de disposer de nouvelles connaissances pour la sélection génétique (partie 3) et d’un référentiel des pratiques alimentaires ou de conduites d’élevage existantes (partie 4) tout en considérant leurs conséquences sur d’autres performances de l’animal ou du système d’élevage. Un éclairage sur la mise en œuvre de pratiques dans des milieux tempérés et pour des zones chaudes à fortes contraintes pédoclimatiques est nécessaire pour envisager des voies d’optimisation ou de transformation des systèmes d’élevage de ruminants (partie 5).
1. Les outils pour phénotyper les émissions de méthane entérique
Les recherches s’appuient sur un ensemble de mesures directes et indirectes afin de progresser à la fois sur les mécanismes biologiques sous-jacents aux émissions de méthane et sur le développement d’outils adaptés aux contraintes de terrain. Les méthodes de mesures directes reposent sur la quantification des flux d’émissions de méthane, tandis que les méthodes d’estimations indirectes reposent sur l’utilisation de modèles de prédiction, de nature diverse. Les variables prédictives peuvent être liées à la ration alimentaire, aux caractéristiques de l’animal (poids vif, niveau de production…) ou être issues de l’analyse de différentes matrices biologiques prélevées sur l’animal (gaz émis, lait, sang, fèces).
1.1. Des mesures directes pour acquérir des données de référence
Jusque-là, différents dispositifs ont permis des mesures directes d’émissions de méthane entérique, et sont utilisés par la plupart des organismes de recherche (Mesgaran et al., 2021) et notamment dans les unités expérimentales d’INRAE : les chambres respiratoires (enceintes dont toutes les entrées et sorties de gaz sont analysées) considérées comme la méthode de référence, l’utilisation de l’hexafluorure de soufre (SF6, un gaz traceur administré par voie orale avec un vecteur qui permet sa libération contrôlée à un flux constant dans le rumen de l'animal), et les GreenFeed® qui permettent d'effectuer des mesures ponctuelles de méthane émis par l’animal lorsqu'il visite un distributeur automatique de concentrés intégré au système. Dans les systèmes d’élevage européens, Doreau et al. (2018) ont montré des niveaux moyens d’émissions de méthane comparables entre les trois outils de mesure pour des lots d’animaux, et validé la fiabilité des mesures avec l’outil GreenFeed® pour une utilisation dans des conditions d’élevage. Cependant, la charge de travail requise pour assurer le fonctionnement des GreenFeed®, leur coût d’acquisition et d’utilisation, et la dépendance à la société qui les commercialise pour l’obtention des données, sont des freins majeurs à leur utilisation en fermes commerciales. Ces dispositifs ont été aussi décrits et discutés pour leur applicabilité en milieu tropical (Gbenou et al., 2024a).
1.2. Des mesures indirectes pour évaluer un grand nombre d’animaux
Des équations de prédiction des émissions de méthane entérique ont été développées depuis plusieurs années afin d’établir des relations entre les émissions mesurées et des facteurs explicatifs tels que les quantités d’aliment ingérées et la composition de la ration pour différentes catégories animales (par exemple, INRA, 2018 ; van Lingen et al., 2019 ; Belanche et al., 2023). Des mesures sur des matrices issues de l’animal (lait et fèces) sont aussi utilisées pour déployer des estimations à plus grande échelle. La composition en acides gras du lait (AGL) a été utilisée comme indicateur des émissions de méthane chez des vaches laitières, initialement nourries avec des aliments à base d’ensilage de maïs contenant des graines de lin extrudées (Chilliard et al., 2009) puis avec une large gamme de régimes alimentaires (Bougouin et al., 2019). La détermination de la composition en AGL se fait par chromatographie en phase gazeuse, ce qui est onéreux et chronophage. Les équations basées sur la composition fine en AGL sont donc peu adaptées à des mesures sur un grand nombre d’animaux. Au contraire, dans le cadre du contrôle de performances, les spectres d’absorption dans le moyen infrarouge (MIR) du lait sont aujourd’hui collectés en routine pour plusieurs millions de vaches afin de prédire la composition globale du lait. Ces spectres ont été mobilisés pour prédire les émissions de méthane chez les bovins laitiers (Vanlierde et al., 2021 ; Fresco et al., 2024b) et permettre une évaluation génomique visant à sélectionner des bovins laitiers faibles émetteurs (partie 3). Des équations de prédiction ont été aussi développées à partir de spectres d’absorption dans le proche infrarouge (SPIR) acquis sur des fèces, une matrice privilégiée pour les femelles allaitantes et les animaux en croissance : des premiers résultats prometteurs ont été obtenus avec un jeu de données de bovins allaitants dont les émissions de méthane ont été mesurées avec des GreenFeed® (Andueza et al., 2022).
Dans tous les cas, l’évaluation de la robustesse des différentes équations de prédiction est nécessaire afin d’estimer avec précision les réductions permises par des stratégies de mitigation déployées à l’échelle du système d’élevage (Benaouda et al., 2019 ; Blondiaux et al., 2024).
1.3. Perspectives de développement des outils de mesure ou de prédicteurs
Une bonne complémentarité entre outils de mesures directes ou indirectes est indispensable si l’on veut aboutir à des prédicteurs suffisamment précis. Les infrastructures expérimentales d’INRAE et de ses partenaires de la recherche et développement s’enrichissent de nouveaux outils complémentaires à ceux déjà utilisés (encadré 2), ce qui nourrira les futurs travaux menés sur les stratégies de mitigation des émissions de méthane entérique dans différents contextes d’élevage. Des chambres d’accumulation mobiles pour petits ruminants (PAC), incluant la mesure durant une heure de gaz émis par flatulence en plus du méthane émis par éructation et expiration (Goopy et al., 2011) sont ainsi disponibles. Le sniffer, positionné au sein d’un robot de traite, permet une mesure par infrarouge de la concentration en méthane émis par le ruminant.
Les conditions et les procédures d’utilisation des méthodes de quantification directe des émissions de méthane entérique par les ruminants ont été consignées dans un ouvrage méthodologique publié par le consortium européen d’infrastructures expérimentales sur les bovins SmartCow coordonné par INRAE (Mesgaran et al., 2021).
Les outils mettant en œuvre ces méthodes sont aujourd’hui déployés dans les stations expérimentales d’INRAE réparties sur le territoire : Le Pin en Normandie (GreenFeed®, sniffers) et l’Installation Expérimentale en Production Laitière de l’UMR Pegase en Bretagne (GreenFeed®) pour les bovins laitiers, Herbipôle en Auvergne pour les bovins laitiers ou allaitants et les ovins (GreenFeed®, chambres respiratoires), le Pôle de Phénotypage des Petits Ruminants en Val de Loire (GreenFeed®) pour les ovins, et la Plateforme Tropicale Élevage pour l’Agroécologie (PTEA) en Guadeloupe (GreenFeed®). Des chambres métaboliques à ambiance contrôlée pour les bovins et des chambres d’accumulation mobiles pour petits ruminants (PAC) seront bientôt disponibles dans différentes stations expérimentales (Herbipôle, Pôle de Phénotypage des Petits Ruminants, unité expérimentale de La Fage dans l’Aveyron, installation expérimentale de l’UMR GenPhySE en Occitanie).
L’Institut de l’Élevage – Idele – a récemment investi dans un parc important de GreenFeed® et un sniffer, qui sont déployables dans les fermes expérimentales laitières, en viande bovine, ou en petits ruminants.
Le potentiel de prédiction de ces méthodes indirectes devra être comparé aux prédictions permises par les spectres MIR du lait, car le développement de prédicteurs adaptés aux contraintes du terrain reste une priorité. En France, des sniffers seront prochainement installés dans 50 fermes commerciales bovines laitières, et des PAC seront utilisées dans différents élevages commerciaux de petits ruminants, pour acquérir des données permettant de comparer les systèmes et d’évaluer les prédicteurs dans des contextes variés.
Des travaux d’amélioration de la précision des prédicteurs déjà identifiés sont également indispensables pour envisager leur utilisation à grande échelle. La complémentarité des recherches conduites sur les bovins et les petits ruminants permettra d’évaluer la possibilité de prédire les émissions de méthane par les spectres MIR du lait ou les SPIR des fèces selon la filière. De nouveaux prédicteurs sont également à l’étude : des métabolites plasmatiques tels que les acides aminés, le glucose, des acides carboxylique ou des glycérolipides (Yanibada et al., 2020), l’abondance isotopique en carbone du lait (Saro et al., 2025), les microorganismes ou les flux d’hydrogène (Bedoya Mazo et al., 2023).
Une acquisition régulière de données adossées à l’expérimentation animale en stations et à des collaborations nationales et internationales, demeure indispensable. L'intégration entre bases de données offrira la possibilité de proposer des combinaisons de modèles encore plus robustes et discriminants pour la prédiction des émissions individuelles de méthane entérique. Cela permettra de tenir compte de la variabilité individuelle dans les évaluations des systèmes d’élevage pour différents contextes pédoclimatiques (partie 5).
2. Le microbiote du rumen : moduler le devenir de l’hydrogène et limiter la production de méthane
Le microbiote ruminal est une cible privilégiée des stratégies de réduction des émissions de méthane, puisque la production de méthane fait suite à la dégradation et la fermentation des aliments par les microorganismes du rumen. Ce sont les archées, un embranchement taxonomique des microorganismes, qui synthétisent le méthane par un ensemble de réactions communément appelées méthanogenèse. Plusieurs voies de méthanogenèse existent mais la plus répandue dans le rumen est la réduction du dioxyde de carbone par l’hydrogène. Ces deux composés ne sont pas produits par les archées mais par les autres microorganismes du rumen : bactéries, protozoaires et champignons (Morgavi et al., 2010). Comme la production de méthane est la conséquence naturelle de la production par le microbiote des substrats énergétiques que sont les AGV pour l’hôte, les stratégies de réduction des émissions de méthane entérique ne doivent pas avoir d’effet négatif sur la fourniture de ces substrats. Plusieurs avancées répondant à ce double enjeu ont été obtenues par INRAE et ses partenaires.
2.1. La gestion de l’hydrogène au sein du microbiote ruminal
La manipulation de la production d’hydrogène et de sa consommation est considérée comme une approche clé pour atténuer les émissions de méthane (Martin et al., 2010 ; Mackie et al., 2024). Les recherches se sont concentrées sur l’évaluation d’accepteurs alternatifs d’hydrogène pour le détourner de la méthanogenèse. En supplémentant la ration de vaches avec des nitrates, un oxydant capable d’être réduit par l’hydrogène, Guyader et al. (2017) ont montré qu’il est possible de stimuler d’autres voies métaboliques acceptant l’hydrogène en compétition avec la méthanogenèse, mais les mécanismes microbiens sous-jacents restent à élucider. Par des approches in vitro, d’autres accepteurs alternatifs d’hydrogène ont été étudiés comme les composés phénoliques (Huang et al., 2023 ; Romero et al., 2023), des métabolites secondaires de plantes que certaines populations microbiennes sont capables de métaboliser en acétate tout en consommant l’hydrogène. Parmi la dizaine de composés phénoliques testés à INRAE, l’acide gallique et le phloroglucinol (ici issus de l’industrie chimique mais qui sont naturellement présents dans les écorces d’arbres) stimulent la production de gaz et d’AGV totaux, notamment l’acétate, sans stimuler la production de méthane. L’effet du phloroglucinol a été évalué en combinaison avec le 2-bromoethanesulfonate, un inhibiteur spécifique des méthanogènes : la production de méthane a été complètement inhibée, l’accumulation d’hydrogène a été réduite comparativement au traitement par le 2-bromoethanesulfonate seul, et la production d’acétate a été augmentée. Les recherches sur la gestion de l’hydrogène continuent à INRAE, visant notamment à l’acquisition de données quantitatives précises sur les transactions d’hydrogène au sein de l’écosystème du rumen.
2.2. Les stratégies dans le jeune âge de l’animal
Après la naissance, le rumen est colonisé de façon séquentielle par le microbiote. Un microbiote est présent dans le rumen dès le deuxième jour de vie et se caractérise par l’abondance des Pseudomonadota (anciennement dénommées protéobactéries), présentes à près de 70 %. La composition évolue rapidement puisque la part des Pseudomonadota chute jusqu’à 30 % avant l’ingestion d’aliments solides, tandis que la part des Bacteroidota, notamment les Prevotella, augmente jusqu’à représenter plus de 50 %. Cette plasticité du microbiote ruminal au début de la vie postnatale de l’animal peut fournir des leviers de réduction des émissions de méthane si les modifications induites persistent à long terme. Chez le veau laitier, l’administration de 3-nitrooxypropanol (3-NOP), un additif inhibiteur de la méthanogenèse, dès la naissance et poursuivie jusqu’à l’âge de 14 semaines, a entraîné une réduction durable de la production de méthane entérique, encore observable à l’âge d’un an (Meale et al., 2021). La composition des bactéries, et dans une moindre mesure celle des archées, a été modifiée dans le rumen, et cette modification persiste avec certaines variations après l’arrêt de la distribution du 3-NOP. En supplémentant la ration de génisses gestantes puis celle de leurs veaux jusqu’au sevrage avec du 3-NOP, d’autres équipes (Martinez-Fernandez et al., 2024) observent une réduction persistante des émissions de méthane 28 semaines après l’arrêt du traitement pour au moins un groupe de veaux traités. Le microbiote ruminal de ces veaux est alors semblable à celui d’une vache adulte faiblement émettrice de méthane mise en évidence dans les troupeaux INRAE de Prim’Holstein (Ramayo‐Caldas et al., 2020).
La modulation du microbiote en début de vie permet de s’affranchir de contraintes qui peuvent être associées à certaines pratiques au sein du système d’élevage comme le pâturage, qui lorsque l’herbe est mature et peu diversifiée, augmente les émissions de méthane chez les ruminants par rapport à un régime à base de concentrés riches en énergie (Roques et al., 2024).
2.3. L’apport de la modélisation mécaniste
La modélisation mécaniste est une approche complémentaire à l’expérimentation pour représenter et prédire le métabolisme ruminal. Un modèle qui représente le microbiote du rumen en groupes fonctionnels associés à l’utilisation du glucose (hexoses), des acides aminés, et de l’hydrogène a été proposé par Muñoz-Tamayo et al. (2016) : il inclut les facteurs d’allocation de flux d’utilisation du glucose et la dynamique de l’hydrogène. Considérer l’hydrogène dans les modèles est capital, mais la connaissance des règles microbiennes et thermodynamiques régissant son métabolisme est encore incomplète, notamment pour la quantification de son devenir et l’impact de l’allocation d’énergie sur la fermentation microbienne (Morgavi et al., 2023). Ce modèle s’avère cependant performant pour représenter des données de fermentation in vitro de régimes mixtes plus ou moins riches en concentrés (Serment et al., 2016). Toutefois, ce modèle ne tient pas compte de conditions physiologiques animales telles que les taux d’absorption des AGV et le passage des substrats. Dans cette optique, l’extension du modèle est en cours pour prendre en compte les conditions in vivo ; les résultats préliminaires comparant les données simulées aux données expérimentales issues de quelques vaches laitières (race Nordic Red) sont prometteurs (Muñoz-Tamayo et al., 2023a). En parallèle, une approche de modélisation a été développée pour relier les données microbiennes à la dynamique des AGV, à l’aide de données provenant d'un système semi-continu in vitro (RUSITEC) imitant certaines conditions in vivo, et de données provenant de rumen de vaches laitières (Davoudkhani et al., 2024).
Le modèle de Muñoz-Tamayo et al. (2016) a été aussi élargi pour représenter l’effet d’une macro-algue (Asparagopsis taxiformis) sur le profil dynamique de fermentation microbienne évalué in vitro : l’inhibition de la production du méthane entraîne une accumulation d’hydrogène qui exerce un contrôle thermodynamique sur la fermentation, via l’allocation du flux vers la production d’AGV. Ce modèle proposé par INRAE (Muñoz-Tamayo et al., 2021) et celui de van Lingen et al. (2021) sont les deux seuls modèles mécanistes permettant de représenter l’effet d’additifs anti-méthanogènes. Au niveau microbien, une approche de modélisation cinétique a été également utilisée pour quantifier les différences métaboliques et énergétiques entre trois espèces de méthanogènes (Muñoz-Tamayo et al., 2019). Les données suggèrent qu’en plus des facteurs cinétiques et thermodynamiques, d’autres forces comme les interactions microbiennes (endosymbiose avec des protozoaires, agrégation microbienne) et la variation spatiale (fractions liquides et particulaires du rumen) contribuent à l’écologie de la communauté des méthanogènes dans le rumen, favorisant la diversité microbienne.
L’intégration des informations génomiques du microbiote doit permettre d’améliorer le pouvoir prédictif des modèles (Muñoz-Tamayo et al., 2023b). En utilisant la séquence génomique de la bactérie cellulolytique Fibrobacter succinogenes S85, il est possible de reconstruire son réseau métabolique et de développer une modélisation métabolique dynamique pour prédire la production d’acétate, de succinate et de formate, à partir du métabolisme du glucose, du cellobiose et de la cellulose (Fakih et al., 2023). Cette approche sera prochainement étendue aux autres microorganismes du rumen, y compris les méthanogènes.
2.4. Perspectives de recherche sur le microbiote
La compréhension des mécanismes de production du méthane par le microbiote permet l’identification de nouvelles stratégies pour réduire les émissions de méthane entérique. Les stratégies de gestion du métabolisme de l’hydrogène ruminal, de modulation du microbiote durant le jeune âge et les approches de modélisation mécaniste pour aborder la complexité microbienne, s’inscrivent toutes dans l’objectif d’identifier les compromis entre réduction des émissions de méthane, valorisation des ressources alimentaires, production, santé et bien-être animal. Deux perspectives de recherche sont identifiées pour poursuivre cet objectif :
i) Élucider les caractéristiques dynamiques de l’écosystème microbien ruminal, notamment sa résilience et les interactions entre espèces microbiennes. Ceci permettrait d’affiner la compréhension du microbiote ruminal et de mieux prendre en compte sa dynamique dans les stratégies de réduction des émissions de méthane entérique ;
ii) Développer un continuum d’études in vivo, in vitro et in silico. Les approches in vitro permettent d’évaluer les voies fermentaires associées aux substrats et aux additifs et les paramètres physico-chimiques associés, pour déconstruire la complexité observée in vivo ; les approches in silico se nourrissent des approches in vivo et in vitro pour prédire l’efficacité des stratégies proposées. Le continuum s’opère dans les deux sens, vers l’in silico pour la compréhension, vers l’in vivo pour l’application. Une telle synergie pour l’élucidation des mécanismes microbiens comme d’ailleurs pour la recherche de proxys contribue aux principes des 3R (Remplacer, Réduire, Raffiner) de l’utilisation d’animaux à des fins scientifiques.
3. Les leviers génétiques pour réduire les émissions de méthane entérique
Pour qu’un caractère soit sélectionnable, il doit être mesurable à relativement grande échelle, et il doit exister une variabilité génétique entre individus. La première condition est assez limitante pour les émissions de méthane qui sont souvent difficilement mesurables au-delà d’un cadre expérimental, c’est-à-dire pour plus de quelques centaines d’animaux. C’est tout l’intérêt des nouvelles techniques de mesure comme les sniffers, mais cette approche n’est pas encore totalement opérationnelle en France. Deux grandes options ont donc été choisies pour réduire les émissions de méthane par la génétique.
3.1. Sélection contre les émissions de méthane prédites à partir des spectres MIR du lait
La prédiction des émissions de méthane à partir des spectres MIR permet d’obtenir de grandes populations d’individus phénotypés, car les spectres sont acquis en très grand nombre ( 15 millions par an pour environ 2,5 millions de vaches laitières) sur plusieurs années, et les prédictions ont un coût marginal faible une fois les formules établies. Des équations ont été développées par deux consortiums de recherche et développement (Optimir et MethaBreed) à partir de mesures de référence obtenues en chambre respiratoire ou par la méthode SF6 (Vanlierde et al., 2021), ou par le GreenFeed® dans neuf conditions expérimentales (Fresco et al., 2024b). Les équations prédisent différents caractères : émission par animal (en g/jour), par unité de production (en g/kg lait standardisé) ou par unité d’ingestion (en g/kg matière sèche ingérée). Une production par jour peut également être reconstituée en multipliant le méthane par kg de lait par la quantité de lait produite. Ces équations ont des précisions modérées. Ce défaut est partiellement compensé par la masse de données qui permet de réduire considérablement la perte de précision au niveau génétique, sans toutefois éliminer les biais.
Ces prédictions ont fait l’objet d’une analyse génétique approfondie (Fresco et al., 2024a), montrant que les héritabilités sont plutôt fortes, ce qui n’est pas surprenant puisque les absorbances des longueurs d’onde les plus informatives d’un spectre MIR sont elles-mêmes relativement héritables. Les corrélations génétiques entre les différents caractères prédits sont modérées, la plus élevée (0,6) étant entre les émissions en g/jour prédites par les équations établies par les deux consortiums. Ce résultat modéré s’explique par des méthodes de mesure et des populations de calibration très différentes entre études des deux consortiums. Les corrélations avec les caractères de production sont également modérées, à l’exception de la prédiction reconstituée, qui est par construction très liée à la quantité de lait ; celle-ci est donc difficilement utilisable car une réduction des émissions de méthane entérique impliquerait une contre-sélection de la production laitière.
Des évaluations génomiques pilotes sont disponibles (Fresco et al., 2024c), réalisées avec un modèle Single Step établi à partir des prédictions à chaque contrôle de performances de plusieurs millions de vaches dont plusieurs centaines de milliers sont génotypées. Cette méthode qui analyse simultanément les performances des animaux typés et non typés estime implicitement l’espérance du génotype des animaux non typés à partir des typages de leurs apparentés, ce qui augmente la précision et diminue les biais. Ces évaluations mettent en évidence l’existence de jeunes taureaux permettant de réduire les émissions de méthane entérique. De plus, elles montrent que les objectifs de sélection récents ont peu influencé le potentiel génétique des animaux quant aux émissions de méthane.
Le déploiement officiel des évaluations génomiques, déterminé par les organismes de sélection, est attendu en 2026. Le caractère méthane choisi est la moyenne des deux prédictions en g/jour (selon les équations obtenues par les deux consortiums), ce qui est plus robuste que le choix d’une seule des deux en l’absence de critère objectif sur l’éventuelle supériorité de l’une ou de l’autre. En l’absence de valorisation économique directe pour l’éleveur, le poids de la réduction des émissions de méthane dans l’objectif global de sélection à définir par les organismes de sélection, peut reposer sur le gain souhaité. Une diminution de 1 % par an des émissions de méthane (en g/jour) impliquerait un poids de l’ordre de 25 % dans l’objectif de sélection (Fresco et al., 2025). Intégrer le méthane aurait pour conséquence un ralentissement modéré du progrès génétique pour les caractères déjà en sélection. Même si aucun effet défavorable n’a été observé jusqu’à présent, il faudra vérifier que la sélection contre le méthane n’a pas d’effet défavorable sur les processus digestifs. Pour les bovins allaitants, l’utilisation des spectres MIR du lait n’est pas envisageable car les vaches ne sont pas traites. Une approche à partir des SPIR sur les fèces est à l’étude. Si elle s’avère efficace, il conviendra de construire un dispositif pérenne de mesure pour envisager ensuite une sélection chez les bovins allaitants.
3.2. Amélioration de caractères d’élevage corrélés aux émissions de méthane
La précision des prédictions MIR est modérée et il existe de nombreux facteurs influençant les émissions de méthane mais qui ne sont pas pris en compte, ou seulement partiellement, dans les prédictions MIR à partir du lait. Il est donc pertinent d’agir aussi sur ces facteurs pour réduire les émissions de manière plus efficace, en complément d’une sélection génétique directe. Il convient de noter que la plupart de ces facteurs dépendent à la fois des systèmes d’élevage et de la génétique, et ce qui est décrit ici vaut autant pour l’orientation de la sélection que pour l’évolution des systèmes d’élevage. En outre, ces améliorations des caractères d’élevage corrélés aux émissions de méthane (âge au vêlage, fertilité, santé, moindre format) ont généralement aussi des effets positifs sur la performance économique de l’élevage (réduction des coûts d’alimentation, des frais vétérinaires…).
Un levier essentiel est de diminuer les pertes, en particulier les mortalités, et les périodes improductives. Cela passe par la réduction du nombre des génisses de renouvellement : plus elles sont nombreuses, plus leur contribution aux émissions de méthane est élevée. Un premier levier est la réduction de l’âge au premier vêlage, qui varie en France de 2 à 3,5 ans avec plus de 65 % du cheptel total (laitier et allaitant) mettant bas à trois ans et plus. Une réduction à deux ans économiserait une cohorte annuelle d’un million d’animaux reproducteurs au format presque adulte, produisant 250 à 300 g de méthane par animal et par jour. Ce changement relève principalement du système d’élevage, mais la sélection sur la précocité peut y contribuer, en particulier pour les races allaitantes généralement plus tardives. L’objectif d’un vêlage à deux ans peut paraître ambitieux et les freins sont divers, mais compte tenu de son très fort impact, tout progrès vers d’avantage de précocité est souhaitable. Chez les bovins allaitants, il convient également de limiter les années sans mise bas, et donc de maximiser la fertilité.
Un deuxième levier est la réduction du taux de renouvellement. Bien que la génétique ne soit pas le facteur le plus important, une amélioration de la longévité par sélection de l’ordre de 0,5 à 1,0 lactation est possible et diminuerait le besoin en génisses de renouvellement. Une réduction du renouvellement à 25 % par an contribuerait à une réduction de 6 % des émissions de méthane du cheptel laitier français dans le cadre d’une première mise bas à deux ans, et jusqu’à 10 % pour des mises-bas à trois ans. Ces résultats sont illustrés par Dall-Orsoletta et al. (2019). Une réduction du taux de renouvellement implique aussi une diminution du taux de réformes involontaires, qu’on peut obtenir par une meilleure santé et une meilleure fertilité des animaux. De plus, les problèmes de santé se traduisent par des pertes de production et donc par des émissions de méthane inutiles. Une sélection sur la santé est possible avec de multiples indicateurs maintenant disponibles : santé de la mamelle ou des pattes, sensibilité à la paratuberculose ou à la cétose ; et la santé et la fertilité sont des caractères avec des poids élevés dans les objectifs de sélection laitiers.
Un troisième levier est la réduction des émissions de méthane relatives aux besoins d’entretien de l’animal, indépendamment de sa production. Le format adulte des races françaises, tant laitières qu’allaitantes, est important. Cela implique une consommation alimentaire élevée tout au long de la carrière. Une réduction de 100 kg du poids adulte, possible par sélection, réduirait les besoins d’entretien de 10 % et induirait une baisse de 3 à 5 % des émissions de méthane selon la part de l’entretien dans l’ensemble des besoins. Elle aurait aussi des effets favorables sur d’autres caractères comme la précocité et la longévité. Cette sélection pour un format plus réduit pourrait s’effectuer sans effet défavorable sur la production laitière car la corrélation génétique entre les deux caractères est très modérée. Une meilleure précocité de développement, facilitée par une réduction de format, favoriserait le dépôt de gras des animaux destinés à la viande, réduisant l’âge à l’abattage et les besoins en concentrés.
Enfin, à système de production donné, les animaux faibles producteurs de lait émettent presque autant de méthane que leurs congénères (Dall-Orsoletta et al., 2019), produisent donc plus de méthane par kg de lait et sont à éviter. La sélection d’animaux avec une meilleure efficacité alimentaire et plus robustes (mieux adaptés à leur milieu) est également un levier.
3.3. Perspectives d’application des leviers génétiques
Les possibilités de réduire l’impact méthane de façon directe et/ou indirecte sont larges. Combiner les différentes possibilités offertes par la génétique est une stratégie développée dans de récents projets collaboratifs comme Méthane 2030 (encadré 3), avec l’objectif de construire un index de synthèse méthane incluant tous les caractères (directs et indirects) associés à des variations de méthane émis. Le succès de cette sélection dépendra avant tout de la pression de sélection réalisée, c’est-à-dire du poids relatif accordé au méthane dans l’objectif de sélection. La pression de sélection ne sera forte que si les éleveurs sont, d’une façon ou d’une autre, incités financièrement à le faire, et donc si les variations d’émissions de méthane induites par la sélection sont mesurables et ont une valeur économique de marché. Des mesures de politique publique ou de filière sont donc nécessaires pour qu’une telle sélection soit mise en œuvre. Il convient de noter que diminuer les émissions de méthane de 1 % par an par la voie d’une sélection directe impose un poids du méthane dans l’objectif de sélection nettement supérieur au prix de marché actuel du carbone. La génétique n’ayant un impact qu’à terme, la valeur à considérer doit refléter la situation dans huit à dix ans au minimum.
4. Principaux leviers alimentaires et de conduite d’élevage pour réduire les émissions de méthane entérique
Les quantités de méthane entérique émises par les ruminants sont donc liées aux micro-organismes du rumen et à la quantité de matière organique à fermenter, mais aussi à la physiologie de l’animal et à sa génétique. Un grand nombre de leviers ont été proposés grâce aux progrès scientifiques et sont aujourd’hui à des niveaux contrastés de maturité, entre découverte scientifique et mise en œuvre à large échelle en fermes commerciales (tableau 1). Le développement de vaccins est en cours dans certains pays en recherchant des antigènes à partir des informations génomiques des méthanogènes du rumen, mais les potentiels effets sur les émissions ne sont pas connus.
Les leviers alimentaires ont un effet modeste (–10 à –20 %) comparativement à certains additifs (–20 à –40 %) mais ont souvent moins de conséquences négatives (observées ou anticipées) sur d’autres composantes de l’animal ou du système (tableau 1). Ils ont un effet immédiat sur la variation des émissions de méthane entérique contrairement aux leviers génétiques où l’effet s’estime à 10 ans. Les variations de l’amplitude des réductions observées avec des leviers alimentaires résultent de la concentration des principes actifs dans les ressources végétales et à de possibles interactions avec d’autres composés de la ration, avec le stade physiologique de l’animal, et(ou) à d’autres facteurs confondants.
La variation d’émissions de méthane entérique constatée suite à la mise en œuvre d’un levier est fonction du mode d’expression choisi : par animal pour les émissions dite « absolues » (en g/jour), par unité d’ingestion pour le « rendement » d’émissions (en g/kg de matière sèche ingérée), ou par unité de produit pour « l’intensité » des émissions (en g/kg lait standardisé ou de viande). Certains modes d’expression (g/jour) sont plus adaptés à l’orientation des politiques publiques et au bénéfice attendu sur le climat, et elles sont donc à privilégier. D’autres (g/kg de produit par exemple) répondent plus directement aux enjeux des éleveurs ou des industries, entraînant potentiellement des conflits entre parties prenantes quant à l’acceptation des leviers.
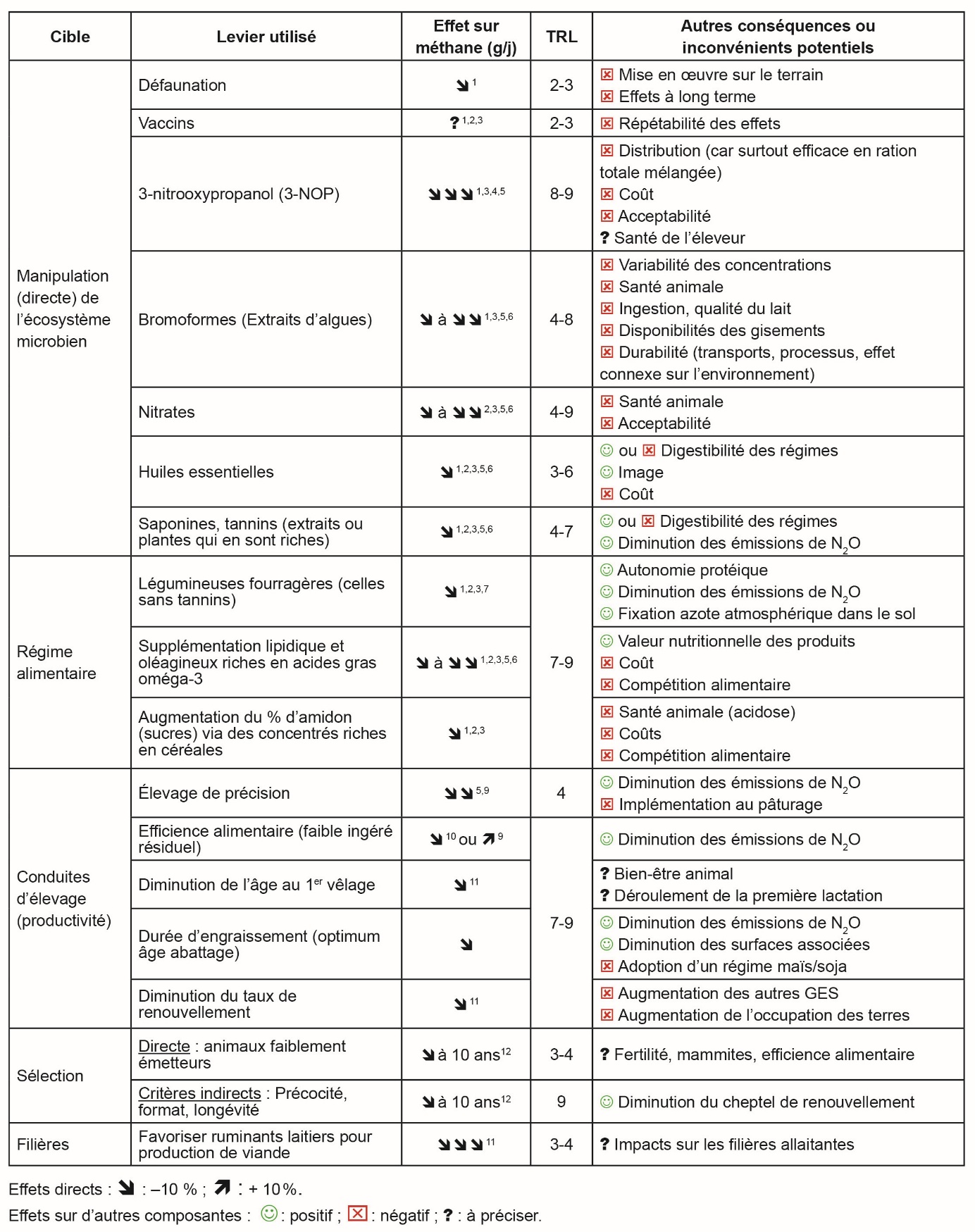
D’après 1 Beauchemin et al. (2022) ; 2 Martin et al. (2010) ; 3 Roques et al. (2024) ; 4 Morgavi et al. (2023) ; 5 Martins et al. (2024) ; 6 Martin et al. (2021) ; 7 Archimède et al. (2011) ; 8 Niderkorn & Jayanegara (2025) ; 9 Fischer et al. (2020) ; 10 Bes et al. (2022) ; 11 Dall-Orsoletta et al. (2019) et 12 de Haas et al. (2021).
TRL (Technology Readiness Level) : échelle de 1 – principes de base – à 9 – éprouvé en environnement opérationnel.
5. Émissions de méthane entérique pour des systèmes d’élevage de ruminants situés dans des zones pédoclimatiques contrastées
5.1. Variabilité des émissions de méthane entérique
Plusieurs pratiques alimentaires (hors sol, pâturage, parcours, agroforesterie, rations mixtes associant fourrages et concentrés) utilisant différentes ressources (coproduits ou résidus de culture, sous-produits industriels, plantes arbustives, fourrages ligneux), coexistent dans les systèmes de production animale, en zones tempérées comme en zones chaudes. Elles conduisent à des émissions de méthane entérique variables. En outre, il existe des différences d’émissions de méthane entre systèmes d’élevage en zone tempérée (Archimède et al., 2011 ; Martin et al., 2021) et celles en zones tropicales (Gbenou et al., 2024b). Les espèces animales élevées dans ces milieux sont différentes, et la connaissance de leur fonctionnement biologique reste hétérogène.
Ainsi, les systèmes d’alimentation animale renseignent bien la valeur des aliments, les caractéristiques des animaux et les pratiques de rationnement en zone tempérée. Par exemple, le dernier modèle de rationnement INRA2018 (INRA, 2018) calcule les émissions de méthane sur la base de la matière organique digestible et inclut le niveau de consommation alimentaire et la proportion de concentrés dans l'alimentation, estimant une diminution du rendement en méthane pour chaque point de pourcentage d’augmentation des concentrés alimentaires. Ces systèmes d’alimentation sont moins bien adaptés aux élevages en zones chaudes. À partir d’une revue de la littérature, Quantin et al. (2025) indiquent par exemple que la quantité de méthane émise en g par kg de matière sèche ingérée est de 23,2 (16,9 à 29,1) pour les vaches laitières et de 27,8 (18,7 à 34,5) pour les génisses en Afrique subsaharienne, contre 21,0 en moyenne en zone tempérée. Elle est de 21,9 (16,8 à 26,5) pour les moutons et 31,7 (de 12,6 à 49,8) pour les chèvres en zone subsaharienne contre 18,0 en moyenne en zone tempérée.
Il existe des équations permettant de prédire à la fois les émissions de méthane et les excrétions azotées pour étudier l’ensemble des rejets polluants liés à l’élevage, mais elles présentent des niveaux d’incertitude variés (Benaouda et al., 2019) et ne sont pas toujours bien adaptées à des ressources riches en composés secondaires bioactifs ou aux additifs qui ont des modes d’action spécifiques. L’incertitude liée aux facteurs d’émissions et le manque de métadonnées contribuent à la mauvaise prise en compte de l’impact environnemental des systèmes d’élevage dans les calculateurs terrains de GES et les inventaires (Eugène et al., 2019 ; IPCC, 2019). Des méthodes existent cependant pour en tenir compte, permettant d’améliorer la sélection des instruments de mesures directes des émissions de GES (Hammond et al., 2016), des proxys (Vanlierde et al., 2024) et des modèles (Blondiaux et al., 2024).
5.2. Reconception de systèmes agricoles et émissions de méthane entérique et d’autres GES
Les stratégies de mitigation des émissions de méthane entérique reposent souvent sur une augmentation concomitante de la productivité, de la longévité et de la reproduction du troupeau, pour réduire les émissions par dilution tout en maintenant la viabilité économique des systèmes d’élevage. En milieu tempéré, les élevages intensifs fortement spécialisés privilégiant la performance technico-économique, peuvent ainsi conduire à des émissions de méthane réduites rapportées au lait ou à la viande. Cependant, ils émettent davantage par unité de surface du fait de la concentration animale, et ils ont un fort impact global négatif sur l’environnement, via leurs fortes émissions azotées dans l’air et dans l’eau, et sur la biodiversité (Chemineau et al., 2025). À l’inverse, la transition agroécologique favorise la biodiversité, améliore le stockage de carbone dans les sols et réduit les émissions de protoxyde d’azote ; cependant, elle a peu d’effets sur la réduction des émissions de méthane (Blaix et al., 2026). Des leviers pour réduire les émissions de méthane entérique pourront être introduits dans ces systèmes, sous réserve qu'ils soient compatibles avec les pratiques et les principes agroécologiques promouvant notamment les interactions entre organismes et leur intégration dans l’écosystème local.
De nombreux travaux ont notamment mis en avant l’intérêt de la polyculture-élevage qui permet une meilleure efficience environnementale, tout en s’appuyant sur des principes agroécologiques associés au bouclage des cycles et à la valorisation de la biodiversité animale et végétale. L’amélioration des pâturages, la complémentation des régimes, les changements d’usage des terres et races de ruminants sont aussi des stratégies déployées dans les zones chaudes du globe (Thornton & Herrero, 2010). Cependant, ces systèmes sont souvent plus dépendants du pâturage naturel. Les éleveurs assurent l’adaptation du bétail à la rareté des ressources grâce à une gestion quotidienne de l’alimentation (recourt aux fourrages ligneux) et à des stratégies de transhumance. Les animaux utilisent des fourrages dont la quantité et la qualité fluctuent au cours des saisons et entre années. Des travaux au Sahel (Assouma et al., 2018 ; Bois et al., 2020) ont évalué l’impact de cette saisonnalité, montrant que l’ingestion de fourrages en saison sèche induit une plus grande formation de méthane in vitro. Des stratégies alimentaires intégrant de la complémentation peuvent réduire les émissions de méthane par kg de matière sèche ingérée, d’autant plus que l’apport en azote est non limitant (Gbenou et al., 2024b).
Les élevages agropastoraux peuvent être des systèmes agroécologiques d’intérêt car, en valorisant les complémentarités et synergies entre une diversité végétale spontanée et cultivée et une diversité animale à l’échelle des individus (comportement et performances) et du troupeau (allotement), ils ont un faible impact environnemental (Vigan et al., 2017). Si leurs caractéristiques les rendent vulnérables aux aléas, elles offrent des capacités d’adaptation et d’atténuation en favorisant l’intégration entre cultures et élevage (voire le polyélevage) avec des actions positives sur le stockage du carbone et la valorisation des déjections. Une analyse des flux de nutriments permet d’améliorer la compréhension des composantes efficientes et celles amenant des pertes concomitantes aux émissions de méthane (Puech & Stark, 2023), afin de proposer des options favorisant les synergies entre carbone et azote. En effet, le bétail joue un rôle crucial dans la distribution spatiale de la matière organique, des éléments nutritifs du sol, et des stocks de carbone. Par exemple, dans les écosystèmes tropicaux, environ 50 % de la matière sèche de fourrages consommée est restituée au sol (Schlecht et al., 2004). Le bilan carbone d’un territoire pastoral peut même être négatif quand on prend en compte l’ensemble des émissions. Dans un écosystème sylvopastoral en climat semi-aride (Assouma et al., 2019), le carbone fixé sur le cycle annuel (matière organique des sols, bois et racines des ligneux) est supérieur aux émissions dues aux animaux et à leurs déjections (fermentation de méthane des animaux d’élevage et des termites, émissions de GES du sol et des eaux, feu de végétation, motopompe de forage).
Ainsi, en s’adaptant favorablement aux milieux contraints dans lesquels ils évoluaient, les systèmes de polyculture-élevage peuvent être des sources d’inspiration pour des stratégies de mitigation à mettre en œuvre en milieu tempéré, notamment dans les zones les plus soumises à évolution sous l’effet du dérèglement climatique.
5.3 Perspectives en recherche et développement
Un premier enjeu est de développer des connaissances et des outils qui permettent de mieux renseigner la diversité animale et végétale des différents milieux dans lesquels s’insèrent les systèmes d’élevage. Il s’agit de calibrer les modèles existants avec des jeux de données spécifiques et/ou de développer des modèles spécifiques au contexte, en fonction des ressources animales et végétales qui y sont présentes.
Un autre enjeu concerne la mobilisation voire l’élaboration d’indicateurs ou de métriques adéquats, pour mieux évaluer les contributions des différents systèmes d’élevages dans toutes les dimensions de la durabilité. Pour cela, il est important de considérer conjointement l’évaluation des émissions de méthane et d’autres GES, avec les aspects du stockage de carbone et de bouclage des cycles de nutriments (azote et phosphore).
Conclusion
Les développements méthodologiques autour de la mesure des émissions de méthane des ruminants, notamment à partir d’analyses MIR sur le lait ou SPIR sur les fèces, permettent aujourd’hui d’évaluer un grand nombre d’animaux dans des conditions diverses. Ils sont de nature à faciliter l’évaluation de l’efficacité de stratégies de réduction des émissions de méthane pour une grande variété de contextes d’élevage avant d’envisager leur transposabilité en fonction des gisements de ressources alimentaires, de la géographie et de la vulnérabilité face au changement climatique propre à chaque territoire. Ces développements associés aux progrès des connaissances ont aussi permis de prototyper des modèles de sélection pour des animaux moins émetteurs. Les progrès associés à toute sélection génétique, y compris génomique, prennent du temps, a minima cinq ans. Au vu de l’urgence climatique, il est donc crucial d’intégrer rapidement ces avancées dans les objectifs de sélection des différentes races, et de combiner approches génétiques directes et indirectes pour maximiser l’impact de la sélection sur la diminution des émissions de méthane par les ruminants.
Les recherches pour identifier les mécanismes biologiques à l’origine des émissions de méthane par l’animal doivent se poursuivre, en particulier celles visant à mieux comprendre les relations entre la diversité microbienne dans le rumen et les phénotypes de l’animal hôte. Ces recherches tireront parti des nouvelles méthodes d’analyses à haut débit pour définir les relations syntrophiques entre les microorganismes, et d’intégration de données dans des modèles de prédiction ou d’évaluation multicritère. Cela permettra d’identifier de nouvelles solutions, basées par exemple sur les flux d’hydrogène, la capture d’électrons ou la promotion de certains consortia de microorganismes, sans pénaliser la valorisation des glucides pariétaux des fourrages par l’animal, préservant ainsi sa production et sa santé.
Le type de ressources alimentaires est un levier important pour réduire les émissions de méthane. Des fourrages adaptés au changement climatique, mais aussi les légumineuses (notamment celles contenant des tannins) et certains coproduits pourraient ainsi être incorporés de manière plus systématique aux rations des ruminants. Pour cela, il s’agira de connaître la valeur nutritive des différentes ressources et leurs effets sur les émissions des différents GES (méthane et protoxyde d’azote), de s’adapter à la variabilité de leur composition selon les aléas climatiques et de considérer leur disponibilité selon le contexte agronomique et les tensions sur leurs différentes valeurs d’usages (production d’énergie par exemple). Grâce aux progrès déjà effectués et aux projets de recherche et développement en cours (encadré 3), les valeurs de ces ressources pour la productivité (lait ou viande) et leurs impacts environnementaux pourront être intégrés dans des outils d’aide à la décision permettant aux différents porteurs d’enjeux d’arbitrer les compromis à des échelles englobantes, dans l’espace (complémentarité au sein du territoire) et dans le temps (au cours des saisons).
Le projet partenarial Méthane 2030 a été lancé en 2024 et pour quatre ans avec l’objectif d’atteindre 30 % de réduction des émissions de méthane entérique en dix ans en France.
Les ambitions sont i) d’harmoniser les dispositifs d’acquisition des données afin de les déployer à grande échelle, ii) d’élaborer des méthodes de prédiction des émissions de méthane adaptées à une diversité de situations d’élevage en France, iii) d’évaluer des solutions nutritionnelles anti-méthane, iv) de mettre en place et déployer des évaluations génomiques sur les émissions de méthane, incluant un index synthétique génétique « efficience méthane », v) d’évaluer la combinaison de différents leviers et de quantifier des émissions de méthane de différents systèmes, et vi) d’élaborer une boite à outils support au conseil pour accompagner la filière. Une trentaine d’essais terrains sont prévus ainsi que des ateliers participatifs.
Le projet Méthane 2030 ambitionne de fournir un service complet à tous les éleveurs bovins pour leur permettre de réduire les émissions de méthane entérique de leur ferme, et ainsi de poursuivre leurs actions de décarbonation. Il réunit 13 partenaires scientifiques et techniques et 15 fermes et sites expérimentaux en vaches laitières et bovins allaitants, impliquant notamment INRAE, APIS-GENE, Idele, les chambres d’agriculture de Bretagne, ELIANCE, FGE, Races de France et plusieurs fermes expérimentales du réseau F@RM XP. Il est financé par l’État dans le cadre du plan France 2030 et est soutenu par les filières au travers d’APIS-GENE.
En France, les derniers chiffres publiés (Citepa, 2025) indiquent que les émissions de GES poursuivent leur trajectoire de réduction entre 2019 et 2023, mais elles restent en dessous des attentes pour respecter le budget carbone actuellement en vigueur ; tous les grands secteurs émetteurs participent à cette réduction et le sous-secteur de l’élevage y contribue via le repli des émissions de méthane et de protoxyde d’azote. Le rythme de réduction des émissions doit donc s’amplifier pour se placer sur une trajectoire compatible avec l’objectif de neutralité carbone en 2050. Pour le méthane et pour les autres GES, il paraît essentiel de modéliser les gains possibles pour différents types de systèmes d’élevage, afin de disposer de chiffres objectivés scientifiquement à mettre en parallèle des scénarios prospectifs sur la place de l’élevage dans les territoires et au regard des stratégies nationales bas carbone. La connaissance incomplète des interactions entre leviers, lorsqu’ils sont combinés au sein d’une stratégie globale au niveau du système, nécessitera des aller-retours entre modélisation et expérimentation.
Lever les freins économiques et sociologiques pour mieux exploiter les opportunités dans les différents territoires selon les atouts et les contraintes propres à chacun, et orienter les politiques publiques restent encore des challenges pour l’acceptabilité des changements à opérer dans les objectifs de sélection et les pratiques d’élevage. Plusieurs chemins d’évolution sont possibles pour reconcevoir les systèmes de production animale, mais doivent intégrer une conception participative des évolutions à engager avec les acteurs concernés. La rémunération des efforts consentis par les éleveurs dépendra des engagements des filières et de politiques publiques volontaristes.
Contribution des auteurs
Florence Gondret et Xavier Fernandez ont plus spécifiquement contribué à l’introduction, à la partie 4 et aux perspectives, et ils ont coordonné l’ensemble du processus d’écriture. Flavie Tortereau, Maguy Eugène, Donato Andueza, Cécile Martin et Solène Fresco ont plus spécifiquement contribué à la partie 1. Simon Roques, Diego Morgavi, Milka Popova, Rafael Muñoz-Tamayo et Valérie Berthelot ont plus spécifiquement contribué à la partie 2. Didier Boichard, Pauline Martin et Solène Fresco ont plus spécifiquement contribué à la partie 3. Maguy Eugène, Cécile Martin, Amandine Lurette et Mohamed Habibou Assouma ont plus spécifiquement contribué aux parties 4 et 5.
Références
- Andueza, D., Picard, F., Pourrat, J., Vanlierde, A., Nozière, P., Cantalapiedra-Hijar, G., Morgavi, D., De la Torre, A., Dehareng, F., Martin, C., & Renand, G. (2022). Near-infrared spectra from faeces as a proxy of enteric methane emissions and intake in beef cattle [Communication]. 73th EAAP International Symposium on Energy and Protein Metabolism and Nutrition (ISEP). Granada, Spain. https://doi.org/10.1016/j.anscip.2022.07.115
- Archimède, H., Eugène, M., Magdeleine, C. M., Boval, M., Martin, C., Morgavi, D. P., Lecomte, P., & Doreau, M. (2011). Comparison of methane production between C3 and C4 grasses and legumes. Animal Feed Science and Technology, 166, 59-64. https://doi.org/10.1016/j.anifeedsci.2011.04.003
- Assouma, M. H., Hiernaux, P., Lecomte, P., Ickowicz, A., Bernoux, M., & Vayssières, J. (2019). Contrasted seasonal balances in a Sahelian pastoral ecosystem result in a neutral annual carbon balance. Journal of Arid Environments, 162, 62-73. https://doi.org/10.1016/j.jaridenv.2018.11.013
- Assouma, M. H., Lecomte, P., Hiernaux, P., Ickowicz, A., Corniaux, C., Decruyenaere, V., Diarra, A. R., & Vayssières, J. (2018). How to better account for livestock diversity and fodder seasonality in assessing the fodder intake of livestock grazing semi-arid sub-Saharan Africa rangelands. Livestock Science, 216, 16-23. https://doi.org/10.1016/j.livsci.2018.07.002
- Beauchemin, K. A., Ungerfeld, E. M., Abdalla, A. L., Alvarez, C., Arndt, C., Becquet, P., Benchaar, C., Berndt, A., Mauricio, R. M., McAllister, T. A., Oyhantçabal, W., Salami, S. A., Shalloo, L., Sun, Y., Tricarico, J., Uwizeye, A., De Camillis, C., Bernoux, M., Robinson, T., & Kebreab, E. (2022). Invited review: Current enteric methane mitigation options. Journal of Dairy Science, 105(12), 9297-9326. https://doi.org/10.3168/jds.2022-22091
- Bedoya Mazo, S., Posada Ochoa, S. L., Rosero Noguera, R., Bayat, A. R., Blondiaux, P., & Eugène, M. (2023). Quantifying H2 emissions under different nutritional mitigation strategies and its impact on improving the prediction of enteric methane emissions of ruminants. Livestock Science, 277, 105342. https://doi.org/10.1016/j.livsci.2023.105342
- Belanche, A., Hristov, A. N., van Lingen, H. J., Denman, S. E., Kebreab, E., Schwarm, A., Kreuzer, M., Niu, M., Eugène, M., Niderkorn, V., Martin, C., Archimède, H., McGee, M., Reynolds, C. K., Crompton, L. A., Bayat, A. R., Yu, Z., Bannink, A., Dijkstra, J., … Yáñez-Ruiz, D. R. (2023). Prediction of enteric methane emissions by sheep using an intercontinental database. Journal of Cleaner Production, 384, 135523. https://doi.org/10.1016/j.jclepro.2022.135523
- Benaouda, M., Martin, C., Li, X., Kebreab, E., Hristov, A. N., Yu, Z., Yáñez-Ruiz, D. R., Reynolds, C. K., Crompton, L. A., Dijkstra, J., Bannink, A., Schwarm, A., Kreuzer, M., McGee, M., Lund, P., Hellwing, A. L. F., Weisbjerg, M. R., Moate, P. J., Bayat, A. R., … Eugène, M. (2019). Evaluation of the performance of existing mathematical models predicting enteric methane emissions from ruminants: Animal categories and dietary mitigation strategies. Animal Feed Science and Technology, 255, 114207. https://doi.org/10.1016/j.anifeedsci.2019.114207
- Bes, A., Nozière, P., Renand, G., Rochette, Y., Guarnido-Lopez, P., Cantalapiedra-Hijar, G., & Martin, C. (2022). Individual methane emissions (and other gas flows) are repeatable and their relationships with feed efficiency are similar across two contrasting diets in growing bulls. Animal, 16(8), 100583. https://doi.org/10.1016/j.animal.2022.100583
- Blaix, C., Dumont, B., Bloor, J. M. G., Zagaria, C., Fleurance, G., Joly, F., & Huguenin-Elie, O. (2026). Agroecological interventions increase biodiversity and the potential for climate change mitigation in Europe. Agriculture, Ecosystems & Environment, 395, 109938. https://doi.org/10.1016/j.agee.2025.109938
- Blondiaux, P., Cabezas-Garcia, E. H., Senga Kiesse, T., Muñoz-Tamayo, R., Reed, K., & Eugène, M. (2024). Evaluation of the precision and accuracy of models of enteric methane emissions from ruminants using Bayesian inference. Zenodo, 140049835. https://doi.org/10.5281/zenodo.14049835
- Bois, B., Morgavi, D. P., González-García, E., Genestoux, L., Lecomte, P., Ickowicz, A., & Doreau, M. (2020). Indirect measures of methane emissions of Sahelian zebu cattle in West Africa, role of environment and management. Tropical Animal Health and Production, 52(4). 1953-1960. https://doi.org/10.1007/s11250-020-02212-x
- Bougouin, A., Appuhamy, J. A. D. R. N., Ferlay, A., Kebreab, E., Martin, C., Moate, P. J., Benchaar, C., Lund, P., & Eugène, M. (2019). Individual milk fatty acids are potential predictors of enteric methane emissions from dairy cows fed a wide range of diets: Approach by meta-analysis. Journal of Dairy Science, 102(11), 10616-10631. https://doi.org/10.3168/jds.2018-15940
- CGAAER. (2024). Parangonnage sur la diminution des émissions de méthane de l’élevage (Rapport no 23065). https://agriculture.gouv.fr/parangonnage-sur-la-diminution-des-emissions-de-methane-de-lelevage
- Chemineau, P., Peyraud, J.-L., Rieu, M., Magdelaine, P., Duru, M., Vermot-Desroches, C., Allo, C., Brulhet, J., & Le Déaut, J.-Y. (2025). L’élevage intensif en France peut-il être durable ? Quelle conciliation entre producteurs, citoyens et consommateurs ? (Controverses de l’Académie d’Agriculture de France 2024). Académie d’Agriculture de France. https://www.academie-agriculture.fr/publications/publications-academie/avis/rapport-de-lacademie-dagriculture-de-france-lelevage
- Chilliard, Y., Martin, C., Rouel, J., & Doreau, M. (2009). Milk fatty acids in dairy cows fed whole crude linseed, extruded linseed, or linseed oil, and their relationship with methane output. Journal of Dairy Science,92(10), 5199-5211. https://doi.org/10.3168/jds.2009-2375
- Citepa. (2025). . Émissions de gaz à effet de serre et de polluants atmosphériques en France 1990-2024 (Rapport Secten éd. 2025). Citepa. https://www.citepa.org/donnees-air-climat/donnees-gaz-a-effet-de-serre/secten/
- Dall-Orsoletta, A. C., Leurent-Colette, S., Launay, F., Ribeiro-Filho, H. M. N., & Delaby, L. (2019). A quantitative description of the effect of breed, first calving age and feeding strategy on dairy systems enteric methane emission. Livestock Science, 224, 87-95. https://doi.org/10.1016/j.livsci.2019.04.015
- Davoudkhani, M., Rubino, F., Creevey, C. J., Ahvenjä Rvi, S., Bayat, A. R., Tapio, I., Belanche, A., & Muñoz-Tamayo, R. (2024). Integrating microbial abundance time series with fermentation dynamics of the rumen microbiome via mathematical modelling. PLoS One, 19(3), e0298930. https://doi.org/10.1371/journal.pone.0298930
- de Haas, Y., Veerkamp, R. F., de Jong, G., & Aldridge, M. N. (2021). Selective breeding as a mitigation tool for methane emissions from dairy cattle. Animal, 15(Suppl 1), 100294. https://doi.org/10.1016/j.animal.2021.100294
- Doreau, M., Arbre, M., Rochette, Y., Lascoux, C., Eugène, M., & Martin, C. (2018). Comparison of 3 methods for estimating enteric methane and carbon dioxide emission in nonlactating cows. Journal of Animal Science, 96(4), 1559-1569. https://doi.org/10.1093/jas/sky033
- EEA. (2024). National emissions reported to the UNFCCC and to the EU under the Governance Regulation. European Environment Agency. https://doi.org/10.2909/6331f651-8863-4656-a911-669f2a332a1e
- Eugène, M., Sauvant, D., Nozière, P., Viallard, D., Oueslati, K., Lherm, M., Mathias, E., & Doreau, M. (2019). A new Tier 3 method to calculate methane emission inventory for ruminants. Journal of Environmental Management, 231, 982-988. https://doi.org/10.1016/j.jenvman.2018.10.086
- Fakih, I., Got, J., Robles-Rodriguez, C. E., Siegel, A., Forano, E., & Muñoz-Tamayo, R. (2023). Dynamic genome-based metabolic modeling of the predominant cellulolytic rumen bacterium Fibrobacter succinogenes S85. mSystems, 8(3), e01027-22. https://doi.org/10.1128/msystems.01027-22
- FAO. (2023). Pathways towards lower emissions – a global assessment of the greenhouse gas emissions and mitigation options from livestock agrifood systems. Food and Agriculture Organization of the United Nations. https://doi.org/10.4060/cc9029en
- Fischer, A., Edouard, N., & Faverdin, P. (2020). Precision feed restriction improves feed and milk efficiencies and reduces methane emissions of less efficient lactating Holstein cows without impairing their performance. Journal of Dairy Science, 103(5), 4408-4422. https://doi.org/10.3168/jds.2019-17654
- Fresco, S., Boichard, D., Aguerre, S., Fritz, S., & Martin, P. (2025). Short communication: First insights into breeding objectives for reducing methane emissions in Montbéliarde dairy cows. Animal, soumis.
- Fresco, S., Boichard, D., Fritz, S., & Martin, P. (2024a). Genetic parameters for methane production, intensity, and yield predicted from milk mid-infrared spectra throughout lactation in Holstein dairy cows. Journal of Dairy Science, 107(12), 11311-11323. https://doi.org/10.3168/jds.2024-25231
- Fresco, S., Vanlierde, A., Baur, A., Boichard, D., Aguerre, S., Fritz, S., & Martin, P. (2024c). Vers une évaluation génomique des émissions de méthane dans les races bovines laitières françaises [Communication poster]. 27e Rencontres Recherches Ruminants, Paris, France. https://journees3r.fr/textes3r/20240903-vers-une-evaluation-genomique-des-emissions-de-methane-dans-les-races-bovines-laitieres-francaises/
- Fresco, S., Vanlierde, A., Boichard, D., Lefebvre, R., Gaborit, M., Bore, R., Fritz, S., Gengler, N., & Martin, P. (2024b). Combining short-term breath measurements to develop methane prediction equations from cow milk mid-infrared spectra. Animal, 18(7), 101200. https://doi.org/10.1016/j.animal.2024.101200
- Gbenou, G. X., Assouma, M. H., Bastianelli, D., Kiendrebeogo, T., Bonnal, L., Zampaligre, N., Bois, B., Sanogo, S., Sib, O., Martin, C., & Dossa, L. H. (2024b). Enteric methane emissions from zebu cattle are influenced by seasonal variations in rangeland fodder quality and intake. Animal, 18(10), 101320. https://doi.org/10.1016/j.animal.2024.101320
- Gbenou, G. X., Assouma, M. H., Zampaligre, N., Martin, C., Bastianelli, D., Bonnal, L., Kiendrebeogo, T., Sib, O., Bois, B., Sanogo, S., & Dossa, L. H. (2024a). Mesurer, prédire et réduire les émissions de méthane entérique en Afrique subsaharienne. INRAE Productions Animales, 37(1), 7648. https://doi.org/10.20870/productions-animales.2024.37.1.7648
- Goopy, J. P., Woodgate, R., Donaldson, A., Robinson, D. L., & Hegarty, R. S. (2011). Validation of a short-term methane measurement using portable static chambers to estimate daily methane production in sheep. Animal Feed Science and Technology, 166(Special Issue: Greenhouse Gases in Animal Agriculture - Finding a Balance between Food and Emissions), 219-226. https://doi.org/10.1016/j.anifeedsci.2011.04.012
- Guyader, J., Ungerfeld, E. M., & Beauchemin, K. A. (2017). Redirection of metabolic hydrogen by inhibiting methanogenesis in the rumen simulation technique (RUSITEC). Frontiers in Microbiology, 8, 393. https://doi.org/10.3389/fmicb.2017.00393
- Hammond, K. J., Crompton, L. A., Bannink, A., Dijkstra, J., Yanez-Ruiz, D. R., O'Kiely, P., Kebreab, E., Eugène, M. A., Yu, Z., Shingfield, K. J., Schwarm, A., Hristov, A. N., & Reynolds, C. K. (2016). Review of current in vivo measurement techniques for quantifying enteric methane emission from ruminants. Animal Feed Science and Technology, 219, 13-30. https://doi.org/10.1016/j.anifeedsci.2016.05.018
- Huang, R., Romero, P., Belanche, A., Ungerfeld, E. M., Yanez-Ruiz, D., Morgavi, D. P., & Popova, M. (2023). Evaluating the effect of phenolic compounds as hydrogen acceptors when ruminal methanogenesis is inhibited in vitro – Part 1. Dairy cows. Animal, 17(5), 100788. https://doi.org/10.1016/j.animal.2023.100788
- INRA. (2018). Alimentation des ruminants (4e ed.). Éditions Quae. https://hal.science/hal-02789908/
- IPCC. (2019). 2019 Refinement of the 2006 IPCC guidelines for national greenhouse gas inventories. Intergovernmental Panel on Climate Change. In CH4 emissions from enteric fermentation, 4, (pp. 1033–1048, 10.3.). Cambridge University Press. https://www.ipcc.ch/report/2019-refinement-to-the-2006-ipcc-guidelines-for-national-greenhouse-gas-inventories/
- IPCC. (2023). Sections. In Core Writing Team, Lee, H. & Romero, J. (Eds.), Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. (pp. 35-115). Intergovernmental Panel on Climate Change. https://www.ipcc.ch/report/ar6/syr/downloads/report/IPCC_AR6_SYR_FullVolume.pdf
- Mackie, R. I., Kim, H., Kim, N. K., & Cann, I. (2024). Invited Review – Hydrogen production and hydrogen utilization in the rumen: Key to mitigating enteric methane production. Animal Bioscience, 37(2), 323-336. https://doi.org/10.5713/ab.23.0294
- Martin, C., Morgavi, D. P., & Doreau, M. (2010). Methane mitigation in ruminants: from microbe to the farm scale. Animal, 4(3), 351-365. https://doi.org/10.1017/S1751731109990620
- Martin, C., Niderkorn, V., Maxin, G., Guyader, J., Eugène, M., & Morgavi, D. P. (2021). The use of plant bioactive compounds to reduce greenhouse gas emissions from farmed ruminants. In R. Baines (Ed.), Reducing greenhouse gas emissions from livestock production (pp. 231-260). Burleigh Dodds Science Publishing. https://hal.inrae.fr/hal-03318574
- Martinez-Fernandez, G., Denman, S. E., Walker, N., Kindermann, M., & McSweeney, C. S. (2024). Programming rumen microbiome development in calves with the anti-methanogenic compound 3-NOP. Animal Microbiome, 6(1), 60. https://doi.org/10.1186/s42523-024-00343-2
- Martins, L. F., Cueva, S. F., Wasson, D. E., Almeida, C. V., Eifert, C., de Ondarza, M. B., Tricarico, J. M., & Hristov, A. N. (2024). Effects of dose, dietary nutrient composition, and supplementation period on the efficacy of methane mitigation strategies in dairy cows: A meta-analysis. Journal of Dairy Science, 107(11), 9289-9308. https://doi.org/10.3168/jds.2024-24783
- Meale, S. J., Popova, M., Saro, C., Martin, C., Bernard, A., Lagree, M., Yáñez-Ruiz, D. R., Boudra, H., Duval, S., & Morgavi, D. P. (2021). Early life dietary intervention in dairy calves results in a long-term reduction in methane emissions. Scientific Reports, 11, 3003. https://doi.org/10.1038/s41598-021-82084-9
- Mesgaran, S. D., Baumont, R., Munksgaard, L., Humphries, D., Kennedy, E., Dijkstra, J., Dewhurst, R., Ferguson, H., Terré, M., & Kuhla, B. (2021). Methods in cattle physiology and behaviour research – recommendations from the SmartCow consortium. Publisso. https://books.publisso.de/de/publisso_gold/publishing/books/overview/53/overview%20chapters
- Morgavi, D. P., Cantalapiedra-Hijar, G., Eugène, M., Martin, C., Nozière, P., Popova, M., Ortigues-Marty, I., Muñoz-Tamayo, R., & Ungerfeld, E. M. (2023). Review: Reducing enteric methane emissions improves energy metabolism in livestock: Is the tenet right? Animal, 17(S3), 100830. https://doi.org/10.1016/j.animal.2023.100830
- Morgavi, D. P., Forano, E., Martin, C., & Newbold, C. J. (2010). Microbial ecosystem and methanogenesis in ruminants. Animal, 4(7), 1024-1036. https://doi.org/10.1017/S1751731110000546
- Muñoz-Tamayo, R., Ahvenjärvi, S., Bayat, A. R., & Tapio, I. (2023a). A dynamic mechanistic model of microbial fermentation and methane production in the cow rumen [Communication]. ADSA Annual Meeting, Ottawa, Canada. https://hal.inrae.fr/hal-04181482v1
- Muñoz-Tamayo, R., Chagas, J. C., Ramin, M., & Krizsan, S. J. (2021). Modelling the impact of the macroalgae Asparagopsis taxiformis on rumen microbial fermentation and methane production. Peer Community Journal, 1, e7. https://doi.org/10.24072/PCJOURNAL.11
- Muñoz-Tamayo, R., Davoudkhani, M., Fakih, I., Robles-Rodriguez, C. E., Rubino, F., Creevey, C. J., & Forano, E. (2023b). Review: Towards the next-generation models of the rumen microbiome for enhancing predictive power and guiding sustainable production strategies. Animal, 17, 100984. https://doi.org/10.1016/j.animal.2023.100984
- Muñoz-Tamayo, R., Giger-Reverdin, S., & Sauvant, D. (2016). Mechanistic modelling of in vitro fermentation and methane production by rumen microbiota. Animal Feed Science and Technology, 220, 1‑21. https://doi.org/10.1016/j.anifeedsci.2016.07.005
- Muñoz-Tamayo, R., Popova, M., Tillier, M., Morgavi, D. P., Morel, J.-P., Fonty, G., & Morel-Desrosiers, N. (2019). Hydrogenotrophic methanogens of the mammalian gut: Functionally similar, thermodynamically different - A modelling approach. Plos One, 14(12), e0226243. https://doi.org/10.1371/journal.pone.0226243
- Niderkorn, V., & Jayanegara, A. (2025). Harnessing bioactive-rich forage species in multi-species grasslands: where do we stand? Challenges and future perspectives [Communication]. 23rd EGF symposium, Reading, United Kingdom. https://hal.inrae.fr/hal-05263461v1
- Poux, X., Allen, M., Cain, M., & Patel, V. (2025). Comportement du méthane dans l’atmosphère et analyse des enjeux techniques du PRG100 et du PRG* - application au secteur des ruminants en France et en Europe (Rapport pour l’OFB). AScA. https://hal.science/hal-05033353v1
- Puech, T., & Stark, F. (2023). Diversification of an integrated crop-livestock system: Agroecological and food production assessment at farm scale. Agriculture, Ecosystems & Environment, 344, 108300. https://doi.org/10.1016/j.agee.2022.108300
- Quantin, P., Akinropo, T., Perveeva, K., Hery Ratovoarisoa, M., Assouma, H., & Eugène, M. (2025). Evaluation of the performance of extant models to accurately predict methane emissions from ruminants in a sub-Saharan African context [Communication]. The 9th International Greenhouse Gas & Animal Agriculture Conference, Nairobi, Kenya.
- Ramayo‐Caldas, Y., Zingaretti, L., Popova, M., Estellé, J., Bernard, A., Pons, N., Bellot, P., Mach, N., Rau, A., Roume, H., Perez‐Enciso, M., Faverdin, P., Edouard, N., Ehrlich, D., Morgavi, D. P., & Renand, G. (2020). Identification of rumen microbial biomarkers linked to methane emission in Holstein dairy cows. Journal of Animal Breeding and Genetics, 137(1), 49‑59. https://doi.org/10.1111/jbg.12427
- Romero, P., Huang, R., Jiménez, E., Palma-Hidalgo, J. M., Ungerfeld, E. M., Popova, M., Morgavi, D. P., Belanche, A., & Yáñez-Ruiz, D. R. (2023). Evaluating the effect of phenolic compounds as hydrogen acceptors when ruminal methanogenesis is inhibited in vitro – Part 2. Dairy goats. Animal, 17(5), 100789. https://doi.org/10.1016/j.animal.2023.100789
- Roques, S., Martinez-Fernandez, G., Ramayo-Caldas, Y., Popova, M., Denman, S., Meale, S. J., & Morgavi, D. P. (2024). Recent advances in enteric methane mitigation and the long road to sustainable ruminant production. Annual Review of Animal Biosciences, 12(1), 321-343. https://doi.org/10.1146/annurev-animal-021022-024931
- Saro, C., Martin, C., Cantalapiedra-Hijar, G., Bouchon, M., Chantelauze, C., & Morgavi, D. (2025). Use of 3-nitrooxypropanol in early lactation dairy cows fed a high forage total mixed ration: effect on enteric methane emissions, performance and milk carbon isotopic signature. Journal of Dairy Science, 109(1), 360-371. https://doi.org/10.3168/jds.2025-26843
- Schlecht, E., Hiernaux, P., Achard, F., & Turner, M. D. (2004). Livestock related nutrient budgets within village territories in western Niger. Nutrient Cycling in Agroecosystems, 68(3), 13. https://doi.org/10.1023/B:FRES.0000019453.19364.70
- Serment, A., Giger-Reverdin, S., Schmidely, P., Dhumez, O., Broudiscou, L. P., & Sauvant, D. (2016). In vitro fermentation of total mixed diets differing in concentrate proportion: Relative effects of inocula and substrates. Journal of the Science of Food and Agriculture, 96(1), 160‑168. https://doi.org/10.1002/jsfa.7076
- Thornton, P. K., & Herrero, M. (2010). Potential for reduced methane and carbon dioxide emissions from livestock and pasture management in the tropics. Proceedings of the National Academy of Sciences, 107(46), 19667-19672. https://doi.org/10.1073/pnas.0912890107
- Vanlierde, A., Dehareng, F., Gengler, N., Froidmont, E., McParland, S., Kreuzer, M., Bell, M., Lund, P., Martin, C., Kuhla, B., & Soyeurt, H. (2021). Improving robustness and accuracy of predicted daily methane emissions of dairy cows using milk mid‐infrared spectra. Journal of the Science of Food and Agriculture, 101(8), 3394–3403. https://doi.org/10.1002/jsfa.10969
- Vanlierde, A., Martin, C., Mertens, A., Lorant, N., Le Gall, L., Morel, I., Renand, G., Rochette, Y., Picard, F., Dehareng, F., & Andueza, D. (2024). Estimation of individual CH4 emissions using fecal near infrared spectra for young, dairy and beef cattle [Communication]. 75th Annual Meeting of the European Federation of Animal Science (EAAP), Florence, Italy. https://docs.eaap.org/boa/2024_Florence_EAAP_Book_Abstracts.pdf
- van Lingen, H. J., Fadel, J. G., Yáñez-Ruiz, D. R., Kindermann, M., & Kebreab, E. (2021). Inhibited methanogenesis in the rumen of cattle: Microbial metabolism in response to supplemental 3-nitrooxypropanol and nitrate. Frontiers in Microbiology, 12(2026), 705613. https://doi.org/10.3389/fmicb.2021.705613
- van Lingen, H. J., Niu, M., Kebreab, E., Valadares Filho, S. C., Rooke, J. A., Duthie, C. A., Schwarm, A., Kreuzer, M., Hynd, P. I., Caetano, M., Eugène, M., Martin, C., McGee, M., O’Kiely, P., Hünerberg, M., McAllister, T. A., Berchielli, T. T., Messana, J. D., Peiren, N., … Hristov, A. N. (2019). Prediction of enteric methane production, yield and intensity of beef cattle using an intercontinental database. Agriculture, Ecosystems & Environment, 283, 106575. https://doi.org/10.1016/j.agee.2019.106575
- Vigan, A., Lasseur, J., Benoit, M., Mouillot, F., Eugène, M., Mansard, L., Vigne, M., Lecomte, P., & Dutilly, C. (2017). Evaluating livestock mobility as a strategy for climate change mitigation: Combining models to address the specificities of pastoral systems. Agriculture, Ecosystems & Environment, 242, 89-101. https://doi.org/10.1016/j.agee.2017.03.020
- Yanibada, B., Hohenester, U., Pétéra, M., Canlet, C., Durand, S., Jourdan, F., Boccard, J., Martin, C., Eugène, M., Morgavi, D. P., & Boudra, H. (2020). Inhibition of enteric methanogenesis in dairy cows induces changes in plasma metabolome highlighting metabolic shifts and potential markers of emission. Scientific Reports, 10(1), 15591. https://doi.org/10.1038/s41598-020-72145-w
Résumé
La réduction des émissions de méthane entérique associées aux systèmes d’élevage de ruminants est l’un des objectifs de lutte contre le changement climatique pour de nombreux pays. Si les mesures directes des émissions de méthane entérique sont limitées dans leur déploiement à un petit nombre d’animaux, des méthodes indirectes de prédiction via le lait ou les fèces peuvent être appliquées à un grand nombre d’animaux dans des contextes variés. Ces approches permettent de développer de nouveaux modèles de sélection génétique ou de conduites d’élevage des petits et gros ruminants associées à des niveaux d’émissions de méthane entériques plus faibles. Les recherches pour comprendre les relations entre la diversité microbienne du rumen et les phénotypes des ruminants contribuent à l’identification de nouveaux leviers pour moduler les populations microbiennes et les flux d’hydrogène dans le rumen, avec l’ambition de réduire les émissions de méthane tout en préservant la production et la santé de l’animal. L’alimentation et la conduite des troupeaux constituent aussi des leviers d’intérêt, mais leurs effets sur la réduction des émissions de méthane entérique sont variables selon les contextes pédoclimatiques, notamment en régions chaudes. Des stratégies combinant différents leviers doivent être identifiées pour réduire efficacement les émissions de méthane sans compromettre la santé de l’animal et les services écosystémiques rendus par leurs systèmes d’élevage.
Pièces jointes
Pas de document complémentaire pour cet articleStatistiques de l'article
 Vues: 3279
Vues: 3279
Téléchargements
 PDF: 400
PDF: 400
 XML: 57
XML: 57
Articles les plus lus par le même auteur ou la même autrice
- Laurianne CANARIO, Nicolas BÉDÈRE, Marc VANDEPUTTE, Didier BOICHARD, Jérôme RAOUL, Catherine LARZUL, Quelles génétiques pour les systèmes d’élevages certifiés en agriculture biologique ? , INRAE Productions Animales: Vol. 37 No 2 (2024): Numéro spécial : L’élevage biologique : conditions et potentiel de développement
- Gonzalo CANTALAPIEDRA-HIJAR, Cécile MARTIN, Donato ANDUEZA, Milka POPOVA, Diego MORGAVI, Abimael ORTIZ-CHURA, Benoit GRAULET, Isabelle CASSAR-MALEK, Muriel BONNET, Anne DE LA TORRE CAPITAN, Gilles RENAND, Sébastien TAUSSAT, Isabelle ORTIGUES-MARTY, Pierre NOZIÈRE, Mécanismes digestifs et métaboliques associés aux différences inter-individuelles de l’efficience alimentaire chez le bovin allaitant , INRAE Productions Animales: Vol. 36 No 3 (2023): Dossier « Efficience alimentaire des bovins allaitants » & Articles hors dossier
- Gérard Xavier GBENOU, Mohamed Habibou ASSOUMA, Nouhoun ZAMPALIGRE, Cécile MARTIN, Denis BASTIANELLI, Laurent BONNAL, Timbilfou KIENDREBEOGO, Ollo SIB, Bérénice BOIS, Souleymane SANOGO, Luc Hippolyte DOSSA, Mesurer, prédire et réduire les émissions de méthane entérique en Afrique subsaharienne , INRAE Productions Animales: Vol. 37 No 1 (2024)
- Didier BOICHARD , C. GROHS, C. DANCHIN-BURGE, A. CAPITAN, Les anomalies génétiques : définition, origine, transmission et évolution, mode d'action , INRAE Productions Animales: Vol. 29 No 5 (2016)
- Mauro COPPA, Bruno MARTIN, Sophie HULIN, Pauline GERBER, Joël GUILLEMIN, Jean-Vincent GAUZENTES, Anne PECOU, Farnaz FAZAELI, Donato ANDUEZA, Authentification de la composition de la ration des vaches laitières par une analyse en spectroscopie visible et infrarouge sur le lait et le fromage , INRAE Productions Animales: Vol. 34 No 2 (2021)
- Pauline MARTIN, Sébastien TAUSSAT, Aurélie VINET, Frédéric LAUNAY, Dominique DOZIAS, David MAUPETIT, Daniel VILLALBA, Nicolas C. FRIGGENS, Gilles RENAND, Précocité, efficience et résilience des femelles bovines allaitantes , INRAE Productions Animales: Vol. 36 No 3 (2023): Dossier « Efficience alimentaire des bovins allaitants » & Articles hors dossier
- Sébastien TAUSSAT, Clément FOSSAERT, Gonzalo CANTALAPIEDRA-HIJAR, Laurent GRIFFON, Pauline MARTIN, Gilles RENAND, Paramètres génétiques de l’efficience alimentaire et faisabilité d’une sélection en population bovine Charolaise , INRAE Productions Animales: Vol. 36 No 3 (2023): Dossier « Efficience alimentaire des bovins allaitants » & Articles hors dossier
- Sébastien TAUSSAT, Gonzalo CANTALAPIEDRA-HIJAR, Jacques AGABRIEL, Gilles RENAND, Sylvie BROUARD, Laurent GRIFFON, Pauline MARTIN, Caractériser les déterminants physiologiques et génétiques de l’efficience alimentaire des bovins allaitants : le programme BEEFALIM 2020 , INRAE Productions Animales: Vol. 36 No 3 (2023): Dossier « Efficience alimentaire des bovins allaitants » & Articles hors dossier
- Amandine LURETTE, S. FRERET, A. CHANVALLON, C. EXPERTON, B. FRAPPAT, J. GATIEN, S. DARTOIS, C. MARTINEAU, C. LE DANVIC, D. RIBAUD, A. FATET, M. PELLICER-RUBIO, La gestion de la reproduction en élevages ovins et caprins, conventionnnels et biologiques : états des lieux, intérêt et acceptabilité de nouveaux outils dans six bassins de production en France , INRAE Productions Animales: Vol. 29 No 3 (2016)