

Précocité, efficience et résilience des femelles bovines allaitantes Dossier : Efficience alimentaire des bovins allaitants
Chapeau
Dans un contexte économique et environnemental où l’élevage est de plus en plus questionné, la précocité, l’efficience alimentaire et la résilience sont présentées comme des leviers d’amélioration de l’efficience globale des élevages. Quelle est la situation de ces trois caractères pour les femelles charolaises et quels sont les liens entre la précocité et l’efficience alimentaire des génisses et la résilience de ces femelles une fois devenues des mères allaitantes ?
Introduction
La filière bovine allaitante d'aujourd'hui doit relever de nombreux défis. Premièrement, dans un contexte où l'aliment acheté à l'extérieur de l'exploitation est de plus en plus cher, les animaux doivent être capables d'assurer leurs différentes fonctions de vie, à savoir la croissance, la survie, la production et la reproduction (Blanc et al., 2006), de manière efficiente, et en valorisant des fourrages plutôt que des concentrés. Deuxièmement, l'efficience, qui est généralement limitée à sa dimension alimentaire (lien entre production et aliments consommés), peut également être vue dans une optique plus large, au travers d'une réduction des périodes de vie improductives, qui aurait des impacts aussi bien économiques qu'environnementaux, notamment par la réduction des gaz à effet de serre émis par unité de production. Et troisièmement, dans le contexte actuel de changement climatique, la vache allaitante pourrait être amenée de plus en plus souvent à devoir faire face à des limitations de la quantité de fourrages disponible à certaines périodes de l'année (Giridhar & Samireddypalle, 2015; Rust, 2019).
Le premier point peut être étudié au travers de l'efficience alimentaire. Celle-ci est définie comme la mise en relation de la production de l'animal et de la quantité d'aliment nécessaire pour obtenir cette production (Cantalapiedra-Hijar et al., 2021). Le nombre élevé (et variable au cours du temps) de processus biologiques impliqués rend ce caractère complexe. Plusieurs critères peuvent être utilisés pour l'estimer, les plus couramment utilisés chez les bovins étant l'ingéré résiduel (abrégée en CMJR pour consommation moyenne journalière résiduelle, ou RFI pour « Residual Feed intake », proposé initialement par Koch et al., 1963) ou l'indice de consommation (souvent plébiscité chez les monogastriques). Quel que soit le critère utilisé, une mesure de l'ingestion individuelle des animaux est nécessaire, mais lourde et coûteuse à mettre en œuvre, ce qui limite fortement le phénotypage de ce caractère.
Pour le deuxième point, la réduction des périodes improductives, la plus longue d'entre elles est généralement celle entre la naissance de l'animal et son premier vêlage. Sur la campagne 2018–2019 par exemple, la moyenne en France de l'âge au premier vêlage des Charolaises était de 36 mois (Bidan et al., 2019). Cet âge plutôt tardif n'est pas spécifique de la race charolaise et est représentatif des principales races allaitantes françaises, ce qui peut constituer un problème de compétitivité face à des races anglo-saxonnes plus précoces avec des âges au premier vêlage proches de 24 mois (Minick Bormann & Wilson, 2010). C'est également un levier important pour réduire les émissions de méthane de l'élevage. Pour ce caractère, les choix de l'éleveur concernant le début de la mise à la reproduction jouent un rôle important, mais les caractéristiques propres, liées à la fertilité de l'animal ne sont pas à négliger, et en particulier l'âge de la puberté. Celui-ci peut être déterminé à l'aide de dosage de la progestérone dans le sang. Des études ont montré qu'il était influencé par différentes causes environnementales telles que la saison ou l'alimentation (Cardoso et al., 2020; Schillo et al., 1992) mais qu'une variabilité génétique existait également entre races, mais aussi intra-race (Perry, 2016).
Le troisième point concerne la capacité des animaux à s'adapter à des périodes de restriction alimentaire. La capacité adaptative peut être définie dans ce cadre comme la réponse de l'animal à cette perturbation, son aptitude à mobiliser ses réserves corporelles pour faire face au déficit énergétique et à maintenir sa capacité à produire et à se reproduire, mais aussi à restaurer ces différentes fonctions après la fin de la perturbation (Blanc et al., 2010). Elle représente une facette pour explorer la résilience des animaux. En effet, la résilience peut être définie comme la capacité des animaux à faire face à une perturbation (vague de chaleur, pathogène…), en tenant compte à la fois de l'intensité avec laquelle l'animal est affecté et de sa capacité à revenir rapidement à l'état initial une fois la perturbation terminée (Poppe et al., 2021). Si les études traitant des animaux dans le cadre d'un challenge de restriction alimentaire sont nombreuses chez les bovins allaitants, elles s'intéressent quasi-exclusivement à des animaux en croissance. Les rares études sur le troupeau reproducteur ont montré que les fonctions biologiques dont le maintien était priorisé par la femelle en restriction étaient différentes d'une race à l'autre (De La Torre et al., 2010; D'Hour et al., 1995). Toutefois ces études portaient sur un faible nombre d'animaux et la réponse adaptative des femelles charolaises à la restriction nécessitait d'être davantage explorée.
Il apparaît donc que la précocité, l’efficience et la résilience sont trois caractères d’une importance croissante pour la durabilité tant économique qu’environnementale de la filière allaitante. Nous nous proposons dans cet article de présenter une synthèse des résultats et nouvelles connaissances apportés par une expérimentation de grande envergure sur ces trois caractères et les liens qui les unissent en race charolaise. Après le descriptif du dispositif expérimental et des méthodes utilisées, les précocités sexuelles et de développement des génisses ainsi que leur efficience alimentaire seront explorées, tant sur des aspects environnementaux que génétiques. La réponse moyenne de la race charolaise à un challenge alimentaire sera ensuite étudiée, ainsi que la variabilité des réponses individuelles. Enfin, l’influence des performances des génisses, et en particulier celle de leur efficience alimentaire, sur leur réponse au challenge alimentaire sera détaillée.
1. Le dispositif expérimental
Le principe de ce programme, dont le déroulement est schématisé figure 1, repose sur l’étude des mêmes femelles à différentes étapes de leur vie. Il s’est déroulé de manière parallèle sur deux Unités Expérimentales INRAE : Bourges (https://doi.org/10.15454/1.5483259352597417E12) et Le Pin (https://doi.org/10.15454/1.5483257052131956E12). Dans un premier temps, le développement corporel et sexuel des femelles a été suivi quand elles étaient génisses. Cette partie de l’expérimentation se terminait par un passage en station de contrôle d’ingestion individuelle pour estimer leur efficience alimentaire. Puis, après leur premier vêlage, les génisses devenues vaches ont participé à un challenge alimentaire où la moitié d’entre elles faisaient l’objet d’une restriction alimentaire pendant la période hivernale afin d’étudier leur capacité à s’adapter à la restriction, et à récupérer une fois celle-ci terminée. Ce challenge était répété pendant trois lactations successives pour les femelles de l’unité du Pin uniquement. L’ensemble des animaux a été traité dans le respect des règles éthiques en vigueur, dans des stations et par du personnel habilité, et l’expérimentation a fait l’objet d’une autorisation référencée sous le numéro APAFIS#14764-2018030610486896 v3.
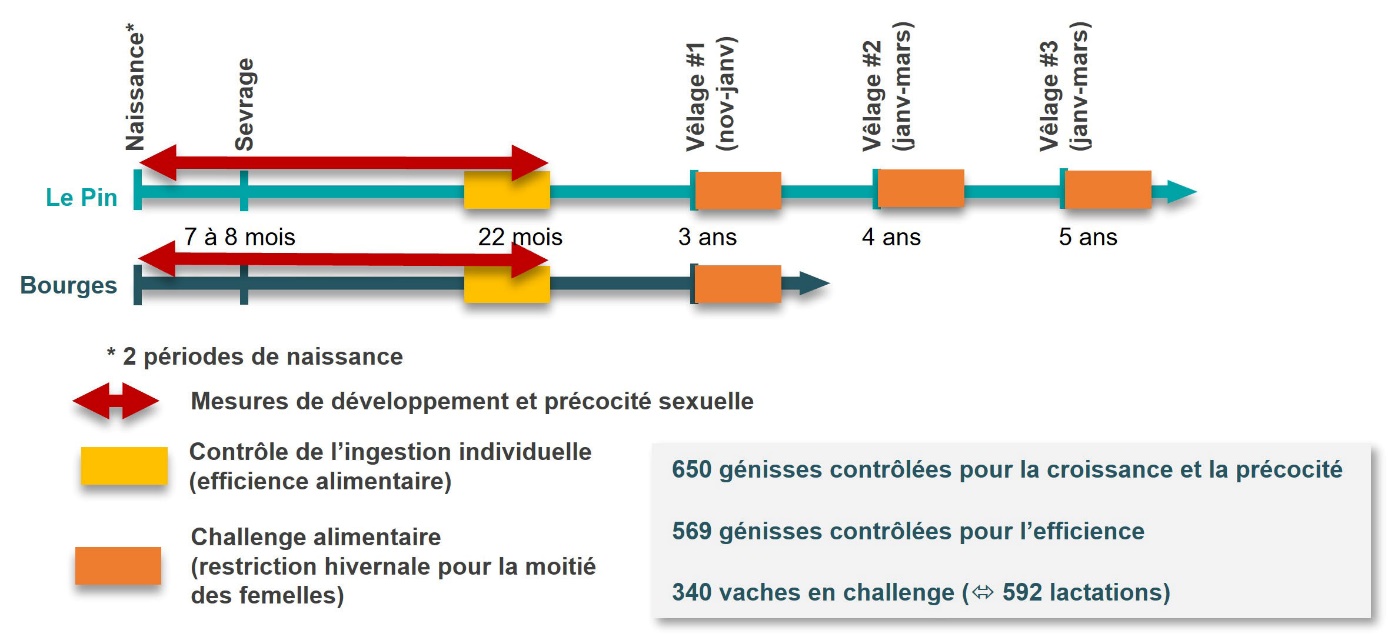
1.1. Croissance, précocité sexuelle et efficience des génisses
a. Procréation des animaux
Entre 2011 et 2015, 650 génisses de race charolaise sont nées sur les deux unités expérimentales INRAE de Bourges (306 animaux) et du Pin (344 animaux). Pour procréer ces animaux, un total de 56 taureaux d’insémination a été utilisé afin de constituer un fond génétique commun entre les deux troupeaux : 48 mâles d’entreprises de sélection (8,5 génisses/père en moyenne) choisis pour leurs caractéristiques extrêmes (améliorateurs ou détériorateurs) en matière d’efficience alimentaire (40 mâles) ou de précocité sexuelle (8 mâles), et 8 mâles eux-mêmes produits sur l’unité du Pin-au-Haras pour favoriser la variabilité en termes de précocité sexuelle (26,0 génisses/père en moyenne). Pour chaque campagne de naissance, les animaux étaient répartis en deux cohortes par unité expérimentale : une avec des naissances en automne – hiver (octobre à début janvier) et une avec des naissances au printemps (fin février à mai).
b. Phénotypes mesurés pendant la croissance
Les animaux ont été pesés à la naissance, puis une fois par mois jusqu’à l’âge de 22 mois. Une double pesée supplémentaire était effectuée au moment du sevrage (entre 7 et 8 mois). Des gains moyens quotidiens (GMQ) pendant différentes périodes et des poids à âge-type ont été déterminés par régression linéaire de ces pesées régulières en fonction du temps. À 18 mois, des mesures de conformations externes (hauteur au garrot, largeur d’épaule, largeur aux trochanters) et internes (pelvimétrie) ont été réalisées.
À partir du sevrage et jusqu'à l'âge de 22 mois, des échantillons de sang ont été prélevés à la queue tous les dix jours afin de déterminer l'âge d'apparition de la puberté par dosage ELISA de la progestérone circulante (Canépa et al., 2008). Les femelles étaient déclarées pubères à partir du moment où deux échantillons successifs présentaient une concentration de progestérone supérieure à 1,5 ng/mL.
c. Évaluation de l’efficience
À 22 mois, l’efficience alimentaire a été mesurée pendant une durée de 12 semaines (après 4 semaines d’adaptation) à l’aide d’auges individuelles équipées de portillons automatiques. Un fourrage grossier (du foin au Pin, complémenté par un kg de concentré et de l’ensilage d’herbe à Bourges) était distribué aux animaux avec pesée quotidienne de l’offert et pesée des refus trois fois par semaine. Le détail des rations est disponible dans l’article introductif de ce dossier. À cette détermination de l’ingestion individuelle s’ajoutaient des pesées toutes les deux semaines. L’efficience alimentaire a été déterminée par le calcul de l’ingéré résiduel (CMJR) obtenu comme le résidu de la régression linéaire suivante :
Où GC est un effet fixe du groupe de contemporains, PMM est le poids moyen métabolique, GMQ est le gain moyen quotidien (régressé intra-ferme), CMJR est l’ingéré résiduel et β1 et β2 sont les coefficients de régression partielle respectifs.
Pour certaines cohortes, un accès à un GreenFeed® (C-Lock, USA) était possible et a permis d'obtenir des mesures d'émissions de méthane entérique pour ces animaux. Le descriptif de ces mesures est consultable dans la publication de Renand et al. (2019).
1.2. Challenge alimentaire des vaches en production
À l'issue de leur passage en station de contrôle de l'ingestion individuelle, les génisses ont été inséminées sur chaleurs naturelles en prévision d'un premier vêlage à 3 ans. Les vêlages ont eu lieu entre fin novembre et février, avec les primipares vêlant en début de saison (jusqu'à début janvier) et les multipares en fin de saison (à partir de mi-janvier). Le calendrier était identique sur les deux unités expérimentales. Dix jours après leur vêlage, les mères entraient dans le challenge alimentaire. Le challenge était divisé en deux phases : i) une phase de restriction « hivernale » qui durait jusqu'à la mise à l'herbe (généralement à la mi-avril) pendant laquelle la moitié des animaux (lot BAS) était nourrie en quantités limitées avec des apports alimentaires inférieurs de 3 UFL à leurs besoins théoriques quand l'autre moitié (lot HAUT) était ad libitum (l'alimentation distribuée pour ce lot représentait 2 UFL de plus par animal que les besoins théoriques) ; et ii) une phase de récupération « printanière », qui s'étendait de la mise à l'herbe jusqu'à mi-juillet, où tous les animaux étaient conduits ensemble sur les mêmes parcelles en un seul lot, en s'assurant de la bonne quantité et qualité de l'herbe à disposition. Le détail des rations distribuées est disponible dans Martin et al. (2022). La période de restriction durait en moyenne 85 jours (écart-type de 28 j), la principale cause de variation étant la date du vêlage. La période de récupération durait, elle, en moyenne 89 jours (écart-type de 11 j) avec une variation principalement liée à la date de mise à l'herbe, adaptée d'une année sur l'autre en fonction des conditions climatiques. Pour tenir compte de ces variations entre individus, les durées de restriction et de récupération ont été incluses en covariables dans les modèles d'analyse.
Les poids des vaches et des veaux, ainsi que les notes d'état corporel des vaches (NEC), étaient déterminés au début et à la fin de chaque période ainsi que toutes les deux semaines. La production laitière pendant 24h était estimée trois fois au cours de la lactation (en milieu et fin de restriction et en milieu de récupération) par pesée du veau avant et après tétées selon la méthode de Le Neindre et al. (1975) validée par Sepchat et al. (2017). Enfin, la reprise de la cyclicité était déterminée par dosage de la progestérone tous les 10 jours à partir de 30 jours après le vêlage jusqu'à l'obtention de deux dosages positifs ( 1,5 ng/mL) successifs (Canépa et al., 2008).
Si l’objectif initial était d’étudier les performances des vaches en production pendant les trois premières lactations, en raison de différentes contraintes pratiques, toutes les cohortes n’ont pas pu être étudiées et seuls des animaux du Pin ont pu effectuer plusieurs lactations. Ainsi, seules 340 femelles ont pu réaliser le challenge sur au moins une lactation (la moitié subissant la restriction et l’autre moitié servant de témoins), pour un total de 592 lactations. Si les femelles réalisaient plusieurs lactations, elles demeuraient dans le même groupe alimentaire (HAUT ou BAS) d’une lactation sur l’autre.
Ce challenge alimentaire visait à étudier la résilience des femelles allaitantes sur le plan de la disponibilité de l’aliment, c’est-à-dire à la fois leur capacité à s’adapter à la perturbation – la restriction alimentaire –, et leur capacité à retourner à leur état initial une fois la perturbation terminée – la récupération.
2. Précocité sexuelle, développement et efficience alimentaire des génisses
2.1. Précocité sexuelle et développement corporel
Environ 10 % des animaux (57 femelles) n'étaient toujours pas cyclés à la fin de la période de prélèvement. Pour ceux dont l'apparition de la cyclicité a eu lieu pendant la période de prélèvement, l'âge moyen de la puberté a été de 16,7 mois (médiane 16,4). Si l'on attribue un âge à la puberté de 22 mois aux femelles qui n'étaient pas encore cyclées, l'âge moyen devient 17,6 mois (médiane 17,0). Ces valeurs sont supérieures à ce qui avait pu être rapporté en race charolaise par le passé, où respectivement 56 % et 60 % des femelles avaient été notées cyclées à 15 mois d'âge (Mialon et al., 1998; Phocas & Sapa, 2004). Une étude récente rapporte également des âges à la puberté moyens plus précoces (autour de 14 mois) chez des animaux issus de mère de différentes races et de père angus ou limousin (Kelly et al., 2022).
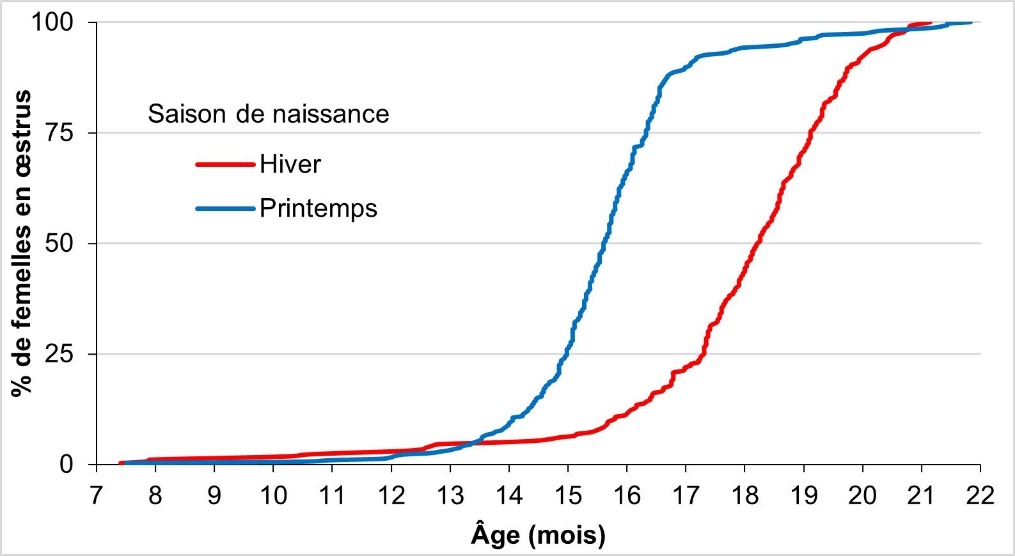
Le lieu d'élevage n'a pas eu d'effet significatif sur l'apparition de la cyclicité. En revanche, l'effet de la saison de naissance a été lui très fort, comme le montre la figure 2 : les femelles nées en hiver (fin octobre à début janvier) étaient pubères plus tardivement que celles nées au printemps (différence de l'ordre de 2 mois et demi pour atteindre 50 % de l'effectif cyclé). Les femelles de printemps (fin février – début mai) présentaient alors des performances comparables à celles de Mialon et al. (1998), d'ailleurs nées entre mi-janvier et mi-avril. Cet effet de la saison sur l'apparition de la puberté a déjà été rapporté dans la littérature (Schillo et al., 1992). La combinaison des variations de durée du jour, de température ambiante et d'exposition à la lumière a été avancée comme explication. Si cette hypothèse manque de travaux récents pour l'étayer, l'identification d'un récepteur de la mélatonine (Tahir et al., 2021) lors d'une analyse d'association tout génome sur l'apparition de la puberté semble aller dans ce sens.
Figure 2. Pourcentage de femelles (génisses charolaises) en œstrus en fonction de l’âge pour chacune des deux saisons de naissance (avec les animaux nés en hiver en rouge et ceux nés au printemps en bleu).
Une héritabilité modérée de 0,21 ± 0,08 a été estimée pour l'âge à la puberté (tableau 1), c'est-à-dire que 21 % de la variabilité observée entre les animaux est d'origine génétique. Les héritabilités rapportées pour ce caractère (à partir de dosages de la progestérone sanguine) dans la littérature sont très variables, avec des estimations allant de 0,07 à 0,67 (Amyes & Morris, 2009; Lefebvre et al., 2021; Martin et al., 1992; Mialon et al., 1998; Morris et al., 2000). Les moyennes se situent généralement autour de 0,4, ce qui est supérieur à ce qui est observé ici. Néanmoins, l'héritabilité estimée est largement suffisante pour envisager une sélection génétique sur ce caractère, qui présente également une certaine variabilité. La difficulté d'une telle sélection réside dans le phénotypage du caractère à grande échelle. Toutefois, les résultats du tableau 1 montrent que d'un point de vue génétique, la précocité sexuelle est corrélée avec des petits poids de naissance et des forts GMQ entre la naissance et le sevrage. Ainsi, même si les valeurs exactes des corrélations nécessiteraient d'être affinées, les erreurs d'estimation étant pour l'instant importantes, une sélection indirecte au travers du poids naissance et de la croissance pré-sevrage peut être envisagée.
Tableau 1. Moyennes, écarts-types, et héritabilités (h²) de l'âge à la puberté et des caractères de poids et de croissance, ainsi que corrélations phénotypiques (Rp), génétiques (Rg) et environnementales (Re) entre l'âge à la puberté et les autres caractères. (Données issues de Taussat et al., 2023, soumis pour publication).
Caractère |
n |
Moyenne (E.T.) |
h² |
Corrélations avec l’âge à la puberté |
||
|---|---|---|---|---|---|---|
Rp |
Rg |
Re |
||||
AP (mois) |
570 |
16,7 (2,2) |
0,21 ± 0,08 |
- |
- |
- |
PDSn (kg) |
650 |
46 (6) |
0,53 ± 0,10 |
– 0,03 |
0,30 ± 0,21 |
– 0,28 ± 0,13 |
GMQs (g/j) |
650 |
957 (173) |
0,25 ± 0,08 |
– 0,28 |
– 0,33 ± 0,23 |
– 0,37 ± 0,08 |
PDSS (kg) |
650 |
265 (43) |
0,24 ± 0,08 |
– 0,34 |
– 0,28 ± 0,25 |
– 0,36 ± 0,08 |
GMQ18 (g/J) |
649 |
564 (150) |
0,26 ± 0,08 |
– 0,13 |
– 0,03 ± 0,27 |
– 0,17 ± 0,08 |
PDS18 (kg) |
649 |
438 (53) |
0,40 ± 0,10 |
– 0,29 |
– 0,23 ± 0,22 |
– 0,35 ± 0,08 |
Signification des caractères : AP= Âge à la puberté (mois), PDSn=Poids naissance (kg), GMQs = Gain moyen quotidien de la naissance au sevrage (g/j), PDSs= Poids au sevrage (kg), GMQ18= Gain moyen quotidien du sevrage à 18 mois (g/j), PDS18= Poids à 18 mois (kg).
Une courbe de Brody (1945) a été utilisée pour modéliser le développement des femelles de l'unité expérimentale du Pin qui effectuaient au moins deux lactations et atteignaient donc leur poids adulte. Pour les 153 femelles considérées, la vitesse de développement, représentée par l'alpha de la courbe de Brody, a été en moyenne de 0,0014 (écart-type 0,0003). Au regard des vitesses de développement rapportées par Lepers et al. (2022), ces animaux ont un développement plus lent que ce qui a été estimé à partir des données nationales pour la race charolaise (0,0016). La corrélation de Pearson (après correction des phénotypes par le groupe de contemporain « année × saison de naissance », la gémellité et l'âge de la mère au vêlage) entre cet alpha de Brody et l'âge à la puberté était de – 0,27 (p 0,0001), ce qui veut dire que les animaux se développant le plus vite sont aussi les plus précoces sexuellement (âge à la puberté plus précoce), sur le plan phénotypique du moins, ce qui est cohérent avec les résultats du tableau 2. Le nombre d’individus n’a pas permis pas l’estimation d’une corrélation génétique.
2.2. Efficience alimentaire des génisses de deux ans
Le passage des animaux en station de contrôle de l'ingestion individuelle a permis d'estimer leur efficience alimentaire autour de l'âge de 2 ans. L'effet de l'unité expérimentale a été important. En effet, les animaux n'ont pas été nourris avec le même type de ration, ceux de Bourges ayant une ration d'ensilage d'herbe tandis que ceux du Pin avaient une ration à base de foin. De ce fait, les génisses de l'unité expérimentale de Bourges ont eu une ingestion légèrement supérieure (+ 0,5 kg/j de MSI) et surtout un GMQ plus important (+ 496 g/j en moyenne). Ces effets ont été corrigés dans les modèles utilisés pour estimer les paramètres génétiques présentés dans le tableau 2. L'héritabilité de la CMJR estimée ici (h² = 0,09) était assez faible par rapport à ce qui est classiquement estimé chez les animaux en croissance, en moyenne autour de 0,3 (Berry & Crowley, 2013), et plus proche de celle estimée chez des animaux à l'entretien, autour de 0,1 (Berry & Crowley, 2013 ; Martin et al., 2019). Cela est probablement lié au faible niveau de croissance des génisses de cet âge et avec cette alimentation, particulièrement au Pin, une forte croissance permettant généralement d'améliorer la précision des modèles (r²) et d'augmenter les valeurs d'héritabilité. Travailler avec des femelles plus jeunes aurait toutefois pu poser un autre souci, car certaines femelles auraient été pubères et d'autres non et cela aurait pu interférer avec la mesure de l'efficience alimentaire.
Si, conformément à la méthodologie employée pour la calculer, la CMJR est phénotypiquement indépendante du GMQ et du poids moyen, on constate néanmoins que génétiquement, les animaux les plus efficients semblent être les plus lourds. Enfin, avec toute la précaution qu'imposent les erreurs standards associées aux estimations des corrélations génétiques en raison de la taille de l'échantillon, les animaux efficients semblent aussi être les plus précoces d'un point de vue génétique. Cette observation est contraire aux rares résultats présents dans la littérature à ce sujet, qui rapportaient plutôt l'absence de corrélation génétique voire une légère corrélation négative entre efficience alimentaire et précocité (Basarab et al., 2011; Crowley et al., 2011; Shaffer et al., 2011). Ces précédentes études se basaient toutefois sur des animaux plus jeunes (entre 10 et 15 mois). Randel et Welsh (2013) ont avancé qu'une estimation de l'efficience réalisée entre le sevrage et la saison de reproduction serait défavorable aux animaux les plus précoces sexuellement et donc avec davantage d'activités de chevauchement. Cela expliquerait la différence observée avec nos résultats qui portent sur des animaux qui ont tous atteint la puberté et pour lesquels il n'y a donc plus cette composante dans l'estimation de la CMJR, qui est à ce stade principalement basée sur le poids métabolique et donc l'entretien.
Tableau 2. Héritabilité (sur la diagonale, en gras, ± erreur standard), corrélations génétiques (au-dessus de la diagonale, en italique, ± erreur standard) et corrélations phénotypiques (sous la diagonale) entre l'efficience, ses composantes et l'âge à la puberté, estimées à partir de 569 génisses phénotypées sur leur efficience, (issu de Taussat et al., 2023, soumis pour publication).
Caractère |
MSI |
GMQ |
PM |
CMJR |
AP |
|---|---|---|---|---|---|
MSI |
0,08 ± 0,07 |
NC |
0,53 ± 0,34 |
0,45 ± 0,42 |
– 0,14 ± 0,44 |
GMQ |
0,33 |
0,00 ± 0,07 |
NC |
NC |
NC |
PM |
0,51 |
0,34 |
0,30 ± 0,10 |
– 0,50 ± 0,35 |
– 0,19 ± 0,26 |
CMJR |
0,84 |
0,00 |
0,02 |
0,09 ± 0,08 |
0,47 ± 0,46 |
AP |
– 0,13 |
0,03 |
– 0,25 |
– 0,03 |
0,21 ± 0,08 |
Signification des caractères : MSI= Matière sèche ingérée, GMQ= Gain moyen quotidien, PM= Poids moyen sur la période, CMJR= Consommation moyenne journalière résiduelle et AP= Âge à la puberté. Pour les corrélations phénotypiques, une valeur absolue supérieure à 0,08 est significative au seuil de 5 %. NC = pas de convergence.
Par ailleurs, les émissions de gaz à effet de serre et les liens phénotypiques entre les niveaux d'émission et l'efficience alimentaire ont été étudiés sur un sous-ensemble (n=326) de génisses de l'expérimentation (Renand et al., 2019). Avec une moyenne de 3,3 mesures par jour et par animal pendant les 12 semaines de contrôle, les variations des émissions de méthane au cours de la journée ont pu être caractérisées : les émissions étaient maximales (autour de 220 g de méthane par jour) en matinée, dans les heures suivant la distribution de l'aliment, puis décroissaient au cours du temps jusqu'à atteindre un minimum de 160 g/j en fin de nuit. Si les émissions de méthane étaient comparables chez les animaux des deux unités expérimentales, les émissions de CO2 étaient, elles, significativement supérieures chez les animaux de Bourges par rapport aux animaux du Pin (6 918 g/j vs 6 139 g/j). Cette différence serait liée au gain de croît supérieur des animaux de Bourges, conséquence de la différence dans les régimes alimentaires des deux unités expérimentales. La corrélation phénotypique entre les émissions des deux gaz était de 0,8, et du même ordre de grandeur que dans de précédentes études chez de jeunes bovins (Alemu et al., 2017 ; Herd et al., 2016 ; Manafiazar et al., 2016) ou des vaches laitières (Pereira et al., 2015). Les émissions de gaz étaient fortement corrélées au poids des génisses (de l'ordre de 0,7 pour le méthane et 0,8 pour le CO2), et modérément avec leur ingestion (entre 0,36 et 0,52 suivant la ferme et le gaz considéré). L'efficience alimentaire (calculée à partir de la CMJR) était indépendante des émissions de méthane lorsque celles-ci étaient exprimées en g/j ou sous forme de résidu d'un modèle linéaire (construit selon le même type de modèle qu'une CMJR). Même si une forte corrélation négative a été observée entre la CMJR et le méthane émis par kg de matière sèche ingérée (MSI) (de l'ordre de – 0,8), l'ensemble de ces résultats confirment la plupart de ce qui est exprimé dans la littérature, à savoir l'indépendance entre efficience et émissions de méthane et qu'il est important de réfléchir en amont à l'unité la plus appropriée suivant l'objectif considéré.
3. Adaptation des vaches face au challenge alimentaire
3.1. Différence entre les groupes restreint et non restreint
a. Influence de la restriction
Les résultats de l'expérimentation ont montré que la restriction alimentaire avait un effet sur l'ensemble des caractères observés sur la période, à savoir le poids de la vache et du veau, les réserves corporelles de la mère, la production laitière et la cyclicité, comme présenté dans le tableau 3. Alors qu'il n'existait aucune différence en termes de poids et d'état corporel entre les deux groupes alimentaires avant l'entrée en challenge, on constate que les animaux non restreints ont pris du poids et de l'état pendant la phase hivernale tandis que les animaux restreints en ont perdu, amenant à une différence entre les deux groupes de 55 kg et 0,55 point de NEC à la fin de la période de restriction. Cette évolution des poids va dans le même sens et est même un peu plus marquée que ce qui avait été observé par De La Torre et al. (2010) dans une première étude chez la race charolaise, qui avait aussi montré une réduction des réserves corporelles révélée par une réduction du diamètre des adipocytes. Une restriction alimentaire modérée (environ – 30 % en énergie et protéines), mais installée dans le temps comme celle que nous avons appliquée, génère un état d'équilibre énergétique négatif chez les animaux. Dans ce cas-là, ils réagissent par des modifications de leur métabolisme qui se traduisent par la mobilisation de leurs réserves corporelles et une diminution de leurs productions (Fruscalso et al., 2013). Petit et Agabriel (1993) avaient montré que chez la vache charolaise, des mobilisations jusqu'à 80 kg de poids vif, similaires à celles de notre étude, étaient constituées à 75 % d'une mobilisation lipidique et à 25 % seulement d'une mobilisation protéique. La baise de NEC observée ici confirme ce point, la NEC étant en effet une estimation du gras sous-cutané qui est un bon indicateur du niveau de réserves lipidiques si la notation est réalisée conformément aux recommandations (Petit & Agabriel, 1993).
Tableau 3. Effets de la restriction alimentaire sur les caractères observés durant le challenge alimentaire (résultats issus de Martin et al., 2022).
Caractère |
Moyenne |
Moyenne |
Différence |
|---|---|---|---|
Poids vache initial (kg) |
699 |
698 |
0,01 |
Poids vache fin restriction (kg) |
712 |
657 |
0,66 |
Poids vache fin récupération (kg) |
740 |
710 |
0,40 |
NEC vache initiale |
2,45 |
2,45 |
0,00 |
NEC vache fin restriction |
2,66 |
2,11 |
0,81 |
NEC vache fin récupération |
2,81 |
2,57 |
0,41 |
Poids veau initial (kg) |
53,6 |
52,9 |
0,08 |
Poids veau fin restriction (kg) |
137,0 |
117,0 |
0,68 |
Poids veau fin récupération (kg) |
239,5 |
216,5 |
0,56 |
Poids veau âge-type 150j (kg) |
200,4 |
179,6 |
0,72 |
Production laitière milieu restriction (kg) |
7,9 |
6,6 |
0,62 |
Production laitière fin restriction (kg) |
7,4 |
5,7 |
0,77 |
Production laitière milieu récupération (kg) |
6,8 |
6,3 |
0,18 |
Reprise de cyclicité* (j) |
105 |
122 |
0,46 |
*Pour la reprise de cyclicité, seule variable pour laquelle il existait une interaction entre rangs de lactation et groupes alimentaires, seules les valeurs corrigées des femelles en première lactation sont présentées ici, les multipares ne présentant aucune différence.
En ce qui concerne la production laitière, il n'était pas possible de comparer le niveau de production des deux groupes alimentaires avant la restriction puisque la première mesure avait lieu en moyenne à 45 jours post-partum soit 35 jours après le début de la phase de restriction. À ce moment, nous avons constaté un écart entre les deux groupes en faveur des femelles nourries ad libitum (7,9 kg vs 6,6 kg), écart qui s'accroissait (7,4 kg vs 5,7 kg) lors de la deuxième mesure, en moyenne 80 jours après le début de la phase de restriction. Une baisse de production laitière dans le cadre d'une restriction alimentaire a largement été rapportée dans la littérature, quel que soit le stade de lactation (Roche, 2007; Tucker et al., 2009), avec des modulations de son intensité notamment selon la race (D'Hour et al., 1995; Gabbi et al., 2016). L'effet sur la production laitière a également rejailli sur les veaux du lot BAS qui pesaient en moyenne 20 kg de moins que ceux du lot HAUT en fin de restriction, alors qu'ils avaient un poids comparable à la naissance. En conséquence, le GMQ des veaux du lot BAS a été plus faible que celui du lot HAUT pendant la phase de restriction (0,8 kg/jour vs 1,0 kg/jour).
Si le rang de lactation avait bien sûr un effet sur le niveau de performance des animaux, l'interaction entre rang de lactation et niveau alimentaire (HAUT ou BAS) n'a pas été significative pour le poids, la NEC, la production laitière et le poids des veaux, ce qui indique que les primipares et les multipares ont répondu de la même manière à la restriction. Cela est contraire à ce qui avait pu être observé chez les races salers et limousine (D'Hour et al., 1995), en lien avec l'hypothèse que les primipares pourraient être plus sensibles à la restriction en raison des besoins énergétiques supplémentaires qu'occasionnent la fin de leur croissance (Freetly et al., 2006). Chez la race charolaise, De La Torre et al. (2010) avaient déjà observé un effet de la restriction plus limité qu'attendu chez les primipares rendant non significatif l'interaction entre le rang de lactation et le régime alimentaire pour certains caractères. Les auteurs expliquaient leurs observations par le niveau de développement particulièrement avancé de leurs animaux primipares, physiquement très proches des multipares. Si la différence de poids entre primipares et multipares était assez marquée dans notre étude, les besoins de croissance des primipares étaient pris en compte dans le calcul de l'alimentation, ce qui peut expliquer que nous ne voyons pas non plus cet effet pour ces caractères.
En revanche, il existe bien une interaction entre régime alimentaire et rang de lactation sur le retour en cyclicité dans notre étude puisqu'une différence entre les groupes alimentaires est observée uniquement chez les primipares (+ 17 j en moyenne pour les primipares du lot BAS). Dans la littérature, différentes études avaient déjà rapporté des effets du rang de lactation (Blanc et al., 2010) ou d'une balance énergétique négative (Butler, 2003) sur la reprise de cyclicité. Dans notre cas, l'interaction observée pourrait bien être le reflet du dispositif expérimental plutôt que d'une différence réelle de capacité entre primipares et multipares face à la restriction. En effet, l'expérimentation a été conduite de façon à ce que les primipares vêlent en début de période de vêlage (fin novembre à début janvier) quand les multipares vêlaient, elles, un peu plus tard (janvier–février). De ce fait, la durée de restriction des primipares était supérieure et seules les primipares étaient encore en phase hivernale au moment de la reprise de cyclicité des femelles du lot HAUT. Pour les multipares des deux lots alimentaires, cela se produisait après la fin de la restriction, ce qui peut expliquer l'absence d'effet observé.
b. Récupération des animaux après restriction
À partir de la mise à l’herbe (généralement mi-avril), les animaux des deux groupes alimentaires étaient conduits ensemble. La restriction était alors terminée et la poursuite des mesures jusqu’à mi-juillet permettait d’observer la récupération des animaux qui avaient été préalablement restreints. En ce qui concerne les vaches, un phénomène de récupération a bien été observé sur l’ensemble des variables puisqu’on constate une diminution des écarts entre les animaux des deux groupes (tableau 3). En revanche, à la fin de la période d’observation, la récupération n’était pas complète puisque ces écarts étaient toujours significatifs. Par exemple, les femelles qui avaient été restreintes pesaient toujours 30 kg de moins et avaient encore un quart de point de NEC de moins que leurs homologues qui n’avaient pas subi de restriction. Toutefois, l’absence de différence entre les animaux des lots HAUT et BAS au vêlage suivant laisse penser que la durée de la période pendant laquelle les mesures ont été effectuées (arrêt mi-juillet) n’a pas été suffisamment longue pour observer la récupération dans son intégralité.
Le début de la récupération des animaux du lot BAS est néanmoins tout à fait observable et il est intéressant de constater que ces femelles montrent une réponse compensatrice, aussi appelée effet rebond (Blanc et al., 2010) à partir du moment où la nourriture redevient abondante. Cela s'exprime notamment au travers du GMQ moyen des femelles du lot BAS plus de deux fois supérieur à celui des femelles du lot HAUT (0,55 kg/j vs 0,25 kg/j). Mais l'expression la plus marquante de cet effet rebond tient dans la reprise de la lactation, avec une production laitière qui augmente après la fin de la restriction (6,3 kg au troisième contrôle, à la moitié de la période de récupération, contre 5,7 kg à la fin de la restriction) alors qu'elle continue de baisser chez les animaux du lot HAUT (6,8 kg contre 7,4 kg). Ces résultats font penser à ce qui peut être observé dans les pays présentant des saisons des pluies très marquées où les courbes de lactation montrent un deuxième pic après le retour de l'abondance alimentaire (Atti, 1998; Koonawootrittriron et al., 2001). L'entrée en phase de récupération a également un effet assez radical sur le retour de la cyclicité puisque la quasi-totalité des animaux qui n'avaient pas encore retrouvé une cyclicité la récupère dans les 15 premiers jours après la mise à l'herbe. Cet effet fort est sans doute lié à une combinaison de facteurs puisqu'en plus du retour de l'abondance alimentaire à l'effet bien connu sur les performances de reproduction (Grimard et al., 2003), l'exposition à la lumière naturelle et l'augmentation de la durée du jour sont aussi connus pour leurs effets positifs sur l'activité ovarienne (Adjorlolo et al., 2019; Yavas & Walton, 2000).
Dans la littérature, l'exemple le plus fréquemment rencontré de réponse adaptative à la restriction est celui de la croissance compensatrice des veaux (Mullins et al., 2021; Pereira et al., 2020). Largement utilisée dans l'industrie de l'engraissement, elle permet des économies d'aliment distribué pour un même niveau de croissance des animaux (Fitzsimmons et al., 2017). Toutefois, dans notre étude, aucune croissance compensatrice n'a été observée chez les veaux issus du lot BAS, qui présentent le même GMQ que ceux du lot HAUT après la mise à l'herbe (1,2 kg/j) et conservent donc une différence de poids d'environ 20 kg. L'absence de croissance compensatrice pourrait être due au jeune âge des animaux au moment de la restriction, des observations anciennes ayant en effet montré que les veaux restreints avant le sevrage ne présentaient en général pas ou peu de croissance compensatrice (Berge, 1991). Ainsi, l'impact de la restriction sur les vaches serait assez limité à long terme, celles-ci récupérant plutôt bien après la fin de la restriction et étant en ce sens résilientes, mais l'absence de récupération des veaux pourrait avoir des conséquences sur le plan économique pour les éleveurs.
c. Priorisation entre fonctions biologiques
Comme nous l'avons vu auparavant, les animaux restreints ont montré des différences de performance pour l'ensemble des caractères étudiés aussi bien en phase de restriction que de récupération. Mais l'importance de l'effet de la restriction alimentaire n'est pas forcément la même pour tous les caractères. Afin de pouvoir comparer ces effets entre eux, il était nécessaire de les exprimer dans la même unité. Pour ce faire, les différences entre les lots HAUT et BAS ont été exprimés en écarts-types du lot HAUT (tableau 3). Il est alors apparu que les vaches charolaises répondaient à la restriction alimentaire de manière plutôt équilibrée avec des effets compris entre 0,66 et 0,80 écart-type pour le poids, la NEC, la production laitière et le poids du veau. En cela, leurs réponses sont intermédiaires entre celles des vaches salers, qui vont privilégier leur production laitière et leur veau aux dépens de leurs propres réserves corporelles, et des limousines, qui vont privilégier leur propre maintien corporel par rapport à celui de leur production laitière (D'Hour et al., 1995). En revanche, la priorisation entre fonctions évolue clairement en faveur de la production laitière au cours de la récupération, la différence entre les deux groupes n'étant plus que de 0,18 écart-type en milieu de récupération par rapport aux 0,4 observés pour le poids et la NEC à la fin de la récupération. L'effet sur la reprise de cyclicité, calculé à partir des primipares seulement, était lui de 0,46 écart-type.
3.2. Variabilité individuelle
Si les résultats présentés dans la partie précédente donnent un éclairage sur la réponse globale des animaux face au challenge alimentaire, on peut également se demander si les vaches sont homogènes dans cette réponse. Pour répondre à cette question, une analyse en composantes principales (ACP) a été réalisée sur les 20 variables décrivant les performances des 293 lactations du groupe restreint (individu statistique = lactation × animal) en termes de poids, de NEC, de production laitière et de reprise de la cyclicité. Cette analyse a été suivie d’une classification hiérarchique sur composantes principales (HCPC) afin d’identifier d’éventuels groupes définissant des profils types de réponse au sein des individus statistiques. Ce travail a été réalisé à partir de résidus des modèles linéaires afin de s’affranchir des effets de l’environnement (ferme × année, rang de lactation, durée de restriction…) et de pouvoir observer la variabilité liée aux individus eux-mêmes.
Figure 3. Résultats de l'analyse en composantes principales présentés sur les dimensions 1 et 2 (A) et 2 et 3 (B) et de la classification des femelles restreintes projetées sur les dimensions 1 et 2 (C), issu de Martin et al. (2023).
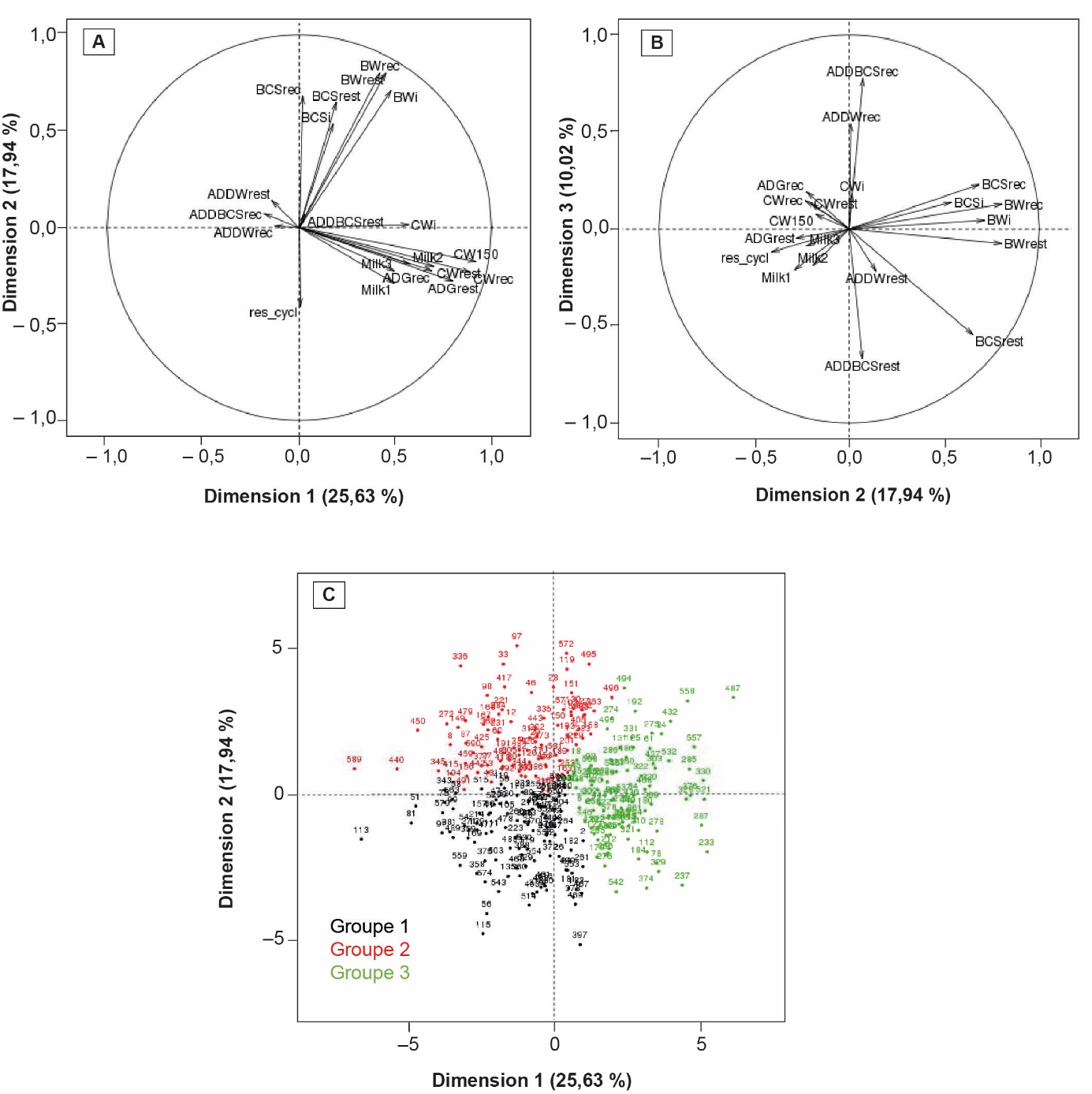
Signification des noms de variables : ADDBCSrec : Différence moyenne quotidienne de NEC durant la récupération, ADDBCSrest : Différence moyenne quotidienne de NEC durant la restriction, ADDWrec : Différence moyenne quotidienne de poids durant la récupération, ADDWrest : Différence moyenne quotidienne de poids durant la restriction, ADGrec: gain moyen quotidien de poids du veau durant la récupération, ADGrest : gain moyen quotidien de poids du veau durant la restriction, BCSi: NEC initiale, BCSrec : NEC à la fin de la récupération, BCSrest : NEC à la fin de la restriction, BWi: poids initial, BWrec : poids à la fin de la récupération, BWrest : poids à la fin de la restriction, CW150 : poids du veau à âge-type 150 jours, CWi : poids initial du veau, CWrec : poids du veau à la fin de la récupération, CWrest : poids du veau à la fin de la restriction, Milk1 : première mesure de la production laitière, Milk2 : deuxième mesure de la production laitière, Milk3 : troisième mesure de la production laitière, et Res_cycl : reprise de la cyclicité ovarienne.
Les résultats de l'ACP (figure 3) montrent que la première dimension, qui explique un quart de la variance totale, est principalement liée aux variables de production laitière et de poids du veau. La deuxième dimension (18 % de variance expliquée) est, elle, davantage liée aux poids des vaches et à leur NEC. La reprise de cyclicité est également projetée majoritairement sur cet axe auquel elle contribue modérément, ce qui illustre la légère corrélation (– 0,2 à – 0,3) existant entre les caractères de poids ou de NEC et la reprise de cyclicité (Martin et al., 2022). Enfin, la troisième dimension (10 % de variance expliquée) est associée aux changements de poids (GMQ) et d'état pendant les phases de restriction et de récupération. La distribution des individus statistiques (animaux × lactation) sur l'ACP forme davantage un continuum que plusieurs groupes distincts, néanmoins, l'analyse HCPC a permis de déterminer une division en trois groupes. Le premier groupe (en noir sur la figure, 111 lactations) contient les animaux avec les plus faibles poids et NEC (aussi bien avant que pendant le challenge), une production laitière et des poids de veau intermédiaires et la plus longue reprise de cyclicité. Le second groupe (en rouge, 89 lactations) contient les animaux avec les poids et NEC les plus importants, une mobilisation de réserves corporelles limitée, la reprise de cyclicité la plus rapide et la production laitière et les poids de veaux les plus faibles. Enfin, le troisième groupe (en vert, 93 lactations) regroupe des animaux de poids et d'état intermédiaires, une reprise de cyclicité longue et les plus fortes productions laitières et poids de veau. Ainsi, au-delà de la réponse moyenne plutôt équilibrée de la race, une certaine variabilité de la réponse existe entre les individus et certains présentent des profils de priorisation plus proches de ce qui avait été décrit par D'Hour et al. (1995) chez la Salers (pour le groupe 3) ou la Limousine (pour le groupe 2). Il est également intéressant de noter que pour les animaux effectuant plusieurs lactations, 57 % d'entre eux demeurent dans le même groupe entre deux lactations successives. Sur trois lactations successives, cette part diminue à 34 %. Cette certaine répétabilité révèle un effet animal qui peut être lié soit à l'environnement permanent, soit à la génétique de l'animal, soit à une combinaison des deux. L'idée que la capacité adaptative ait un contrôle partiellement génétique nécessiterait d'être explorée par une étude spécifique, ce que ne permettent pas les effectifs des animaux restreints de cette expérimentation.
4. Mise en relation des caractères sur l’ensemble de la vie
4.1. À partir des données zootechniques
a. Impact de l’efficience alimentaire des génisses sur leurs performances adultes
Sur la base de l'efficience alimentaire mesurée chez les génisses, deux classes extrêmes de CMJR ont été définies : les femelles efficientes, avec une CMJR inférieure à – 0,4 kg de MSI/j, et les femelles inefficientes, avec une CMJR supérieure à 0,4 kg de MSI/j. Parmi les 592 lactations qui ont été analysées pour le challenge alimentaire, 232 sont issues d'animaux appartenant à l'un de ces deux extrêmes : 115 lactations de femelles efficientes et 117 lactations de femelles inefficientes. Il est tout d'abord intéressant de constater que la classe de CMJR n'a pas d'influence sur la probabilité d'une génisse d'atteindre le premier vêlage (p-value d'un Chi² = 0,70) ni sur celle d'effectuer trois lactations (p-value = 0,34). L'effet de la classe de CMJR et de sa possible interaction avec le régime alimentaire a ensuite été évalué sur 14 caractères mesurés pendant le challenge alimentaire (poids et NEC de la mère et poids du veau en début challenge, en fin de restriction, en fin de récupération, poids à âge-type 150j du veau, les trois productions laitières et le retour de la cyclicité) à l'aide du modèle linéaire décrit et utilisé dans Martin et al. (2022). Il ressort de cette analyse que les génisses efficientes conduisent à des vaches légèrement plus lourdes au moment de leur premier vêlage (+ 18 kg) que les génisses inefficientes. Cette différence augmente même jusqu'à 35 kg au cours du challenge chez les animaux nourris ad libitum mais disparaît chez ceux en restriction. Aucun effet sur la NEC n'a été constaté. En revanche, la production laitière des femelles efficientes est plus faible que celle des femelles inefficientes, avec une différence de – 0,4 kg/j lors du premier contrôle, mi- restriction puis – 0,7 kg/j aux deux autres contrôles, en fin de restriction et en fin de récupération. Cette différence se répercute sur le poids des veaux et est encore plus accentuée chez les veaux du lot BAS où elle se cumule avec l'effet de la restriction alimentaire. Enfin, une interaction entre le groupe alimentaire et la classe de CMJR a été observée sur le retour de la cyclicité. En effet, si le retour en cyclicité des femelles inefficientes n'est pas influencé par le niveau alimentaire, ce n'est pas le cas des femelles efficientes. Ces dernières retrouvent leur cyclicité une semaine plus tôt que les inefficientes quand la nourriture est abondante mais une semaine plus tard que les femelles inefficientes dans le cas d'une restriction alimentaire.
Les études mettant en relation l'efficience des jeunes femelles avec leurs performances ultérieures sont assez rares en races allaitantes. Parmi celles de notre connaissance (Black et al., 2013; Callum et al., 2019; Hafla et al., 2013; Parsons et al., 2021), aucune n'étudie en plus l'impact d'une restriction alimentaire. Ces études concluent généralement à un effet limité de l'efficience de la génisse sur les performances ultérieures, l'un des rares paramètres affectés ultérieurement étant les quantités ingérées, plus élevées chez les femelles inefficientes. La seule étude ayant analysé la production laitière ne constatait pas de différence basée sur l'efficience de la génisse (Black et al., 2013), contrairement à ce que nous avons observé. La méthode pour évaluer la production laitière n'était toutefois pas la même (double pesée du veau vs salle de traite). De plus, le lait était également corrigé pour sa composition, ce qui ne pouvait pas être fait par la technique de la double pesée. Ces éléments peuvent expliquer la différence de résultat. Aucune de ces études ne mesurait le retour en cyclicité après vêlage.
b. Impact de la précocité sexuelle et du développement sur les performances adultes
Afin d’étudier l’impact de la précocité sexuelle et de la vitesse de développement des génisses sur les performances adultes, le même type de modèle linéaire que pour l’étude de l’influence de la CMJR a été utilisé. Ainsi, les animaux ont été séparés en trois classes sur la base de leur précocité sexuelle (précoce : apparition de la cyclicité à 16 mois ou moins, neutre : apparition de la cyclicité à 17 ou 18 mois, et tardive : apparition de la cyclicité à partir de 19 mois), ainsi que sur leur vitesse de développement (lent : alpha0,0013, rapide : alpha0,0015, et neutre pour les valeurs intermédiaires). Les résultats (non publiés par ailleurs) montrent que les animaux les plus précoces sexuellement sont généralement plus légers au début du challenge alimentaire et que cette différence s’estompe progressivement, en particulier chez les animaux nourris ad libitum (tendance uniquement sur le GMQ pendant la phase de restriction). La NEC n’est pas affectée. La production laitière des femelles précoces est en revanche légèrement inférieure à celle des femelles tardives, ce qui a pour conséquence que les veaux des femelles précoces sont plus légers de quelques kilos. Il est néanmoins possible que cet effet soit davantage dû à l’âge de la mère plutôt qu’à sa précocité. En effet, le protocole présente deux saisons de naissance pour les mères et une seule pour les veaux. En raison de l’influence de la saison de naissance sur la précocité, les femelles les plus précoces sont également les plus jeunes au moment de la première mise bas.
Les modèles linéaires n'ont pas permis d'observer un lien entre l'âge à la puberté et le retour en cyclicité. En revanche, si on effectue une corrélation de Pearson entre les valeurs brutes, il s'agit de la seule variable pour laquelle la corrélation est significative, avec les femelles précoces qui reviennent plus tôt en cycle, conformément à ce qui a été observé chez les vaches laitières (Lefebvre et al., 2021).
En ce qui concerne la précocité de développement, les femelles au développement le plus rapide produisent plus de lait que celles au développement tardif, et leurs veaux sont de fait plus lourds.
Contrairement à ce qui a été observé pour la CMJR, il n’y a ici pas d’interaction entre les deux variables de précocité et le niveau alimentaire (hormis la tendance évoquée précédemment pour la précocité sexuelle).
4.2. Au travers d’une modélisation
De manière additionnelle à l'étude directe des performances, les données de l'unité expérimentale du Pin ont également servi à calibrer un modèle de simulation, afin de s'intéresser aux interactions entre variables à l'échelle de la vie entière. Le modèle utilisé est une adaptation du modèle AQAL (AcQuisition et ALlocation de l'énergie) proposé chez la vache laitière par Puillet et al. (2016), et utilise les caractéristiques de la dynamique de troupeau allaitant présentés dans Villalba et al. (2006, 2010). Une classification effectuée sur les performances des génisses au cours de leur période de contrôle de la CMJR a permis de mettre en évidence quatre classes dont deux montraient des CMJR moyennes bien distinctes. Le modèle indique que sur cette période de leur vie, les animaux de la classe la plus efficiente présentent une digestibilité apparente de deux points de pourcentage supérieure à celle de la classe la plus inefficiente, confirmant ce qui a été observé sur des données réelles d'animaux plus âgés par De La Torre et al. (2019). Toujours d'après la simulation, les animaux les plus efficients pendant la période de calcul de la CMJR étaient également plus légers et ingéraient moins que ceux du groupe plus inefficient. Les performances simulées sur l'ensemble de la vie des animaux de ces deux classes ont ensuite été comparées. Déjà plus lourdes pendant la croissance, les animaux de la classe la plus inefficiente deviennent, d'après ces simulations des vaches plus lourdes et plus grasses que celles de la classe efficiente, qu'il y ait ou non restriction alimentaire. Cela ne correspond pas aux performances réelles des animaux, où i) les femelles efficientes sont légèrement plus lourdes que les inefficientes en période d'abondance ; ii) aucune différence n'est constatée pendant la restriction alimentaire ; et iii) aucune différence significative de note d'état n'est observée non plus, quel que soit le contexte alimentaire.
Concernant le retour de la cyclicité, on observe là aussi une différence entre données simulées et données réelles. En effet, quand la ressource alimentaire est abondante, il n'y a pas de différence sur le retour de la cyclicité des deux classes d'après les données simulées alors que sur les données réelles, les efficientes reviennent en cyclicité plus rapidement. Dans le cadre d'une restriction, des animaux simulés de la classe inefficiente ont une NEC qui, cumulée à la perte naturelle post-vêlage, passe sous un seuil de 2,5 utilisé dans les paramètres de calibration du modèle comme prolongeant l'anœstrus post-partum (Sanz et al., 2004). De ce fait, un léger retard apparait dans la simulation sur les aspects reproductifs des femelles inefficientes en restriction, retard qui s'accentue au fur et à mesure des lactations, alors qu'aucune différence de NEC n'est observée sur les données réelles où les animaux inefficients du groupe restreint reviennent plus tôt en cyclicité que leurs homologues efficientes. L'absence de compétition entre fonctions biologiques quand les conditions environnementales sont favorables et l'apparition de compromis quand elles se détériorent, phénomène mis en avant ici par la simulation et les données réelles des femelles efficientes, a également été observé par Douhard et al. (2014) dans un modèle de simulation chez la chèvre laitière.
Concernant la production laitière, celle-ci est supérieure chez les femelles de la classe la moins efficiente, de même que les poids des veaux au sevrage, et ce dans les deux environnements alimentaires, ce qui est cette fois-ci en accord avec les données observées. Cette production supérieure se fait toutefois au prix d’une consommation alimentaire également supérieure.
Le fait de coupler des données expérimentales avec un modèle pour explorer les conséquences sur le long-terme nous paraît une approche prometteuse pour identifier des leviers permettant d’améliorer l’efficience durable de l’élevage allaitant, selon différents scénarios de changement climatique par exemple, bien que quelques améliorations soient encore nécessaires pour être plus fidèle à la réalité. Il est toutefois important de mentionner que dans le cas présent, certains paramètres de calibration du modèle n’ont pas pu être basés sur les données des femelles du projet, puisque le modèle requérait des informations qui n’étaient pas disponibles dans l’expérimentation. La calibration a donc été affinée à partir d’autres jeux de données et de la littérature. Cela peut contribuer à expliquer les différences observées entre la simulation et les données réelles.
Conclusion
L’ensemble des résultats de cette expérimentation, dont certains doivent encore être affinés, plaident pour une prise en compte globale des performances des animaux au cours de leur vie pour l’étude des caractères complexes tels que l’efficience ou la résilience et leur intégration dans des objectifs de sélection. La figure 4 synthétise les principaux résultats de l’expérimentation. La précocité sexuelle apparaît comme un levier particulièrement prometteur pour réduire les périodes improductives à l’échelle de la vie des animaux. Les liens a priori favorables avec la vitesse de développement et l’efficience alimentaire ne font qu’accroître son intérêt. Si le caractère « âge à la puberté » possède en lui-même une héritabilité et une variabilité génétique qui permettraient une sélection, la difficulté tient surtout dans un phénotypage à grande échelle impossible à déployer aujourd’hui. De ce fait, il est nécessaire d’utiliser d’autres caractères et les résultats de notre étude plaident pour une possible sélection pour des petits poids de naissance, utilisée conjointement avec des périodes de naissance de printemps et une alimentation permettant un développement suffisant. D’autres projets de recherche récents ou à venir poursuivent l’exploration de cette question.
Figure 4. Présentation schématique des principaux résultats de l’expérimentation.

L’étude de l’efficience alimentaire soulève la question de l’évolution du classement des animaux d’un stade de la vie à l’autre. Si une évaluation à l’âge de deux ans comme réalisée ici entraîne une corrélation positive entre efficience et âge à la puberté et une indépendance entre efficience et émissions de méthane, la faible croissance des animaux sur la période considérée limite l’héritabilité du caractère. De plus, une sélection sur cette mesure d’efficience alimentaire pourrait avoir des effets indésirables sur les performances des femelles en lactation, en particulier quand les conditions environnementales ne sont pas favorables. Le fait que les résultats puissent être différents si l'efficience était mesurée sur un autre stade de la vie des animaux (et en particulier une période de pleine croissance) jette un doute certain sur l’utilisation qui peut être faite des mesures d’efficience alimentaire et l’impact d’une sélection sur ce caractère.
Enfin, la réponse des femelles allaitantes au challenge alimentaire a montré que les femelles réorganisaient l’allocation de leurs ressources entre les différentes fonctions biologiques dans le cas d’une restriction, mais étaient capables d’un rebond de production au retour de l’abondance alimentaire et d’une récupération complète de leur développement corporel avec le temps, ce qui permet d’évaluer la résilience des femelles charolaises dans un contexte de manque alimentaire, ainsi que la variabilité existante entre individus. L’évaluation de la résilience est néanmoins liée à la nature de la perturbation et on peut se demander également si les animaux les plus résilients ici le seraient aussi dans le cadre d’une perturbation liée au changement climatique (vague de chaleur) ou face à un pathogène.
Remerciements
Les auteurs remercient l’ensemble des personnels des unités expérimentales de Bourges-la-Sapinière et du Pin-au-Haras pour l’ensemble des données collectées pour cette expérimentation. Ils remercient également le personnel du laboratoire de Phénotypage Endocrinologie de Nouzilly pour les dosages de progestérone.
Ce travail a bénéficié du soutien financier du fond pour la recherche et l’innovation Horizon 2020 de l’Union Européenne (projet GenTORE, no 727213) et du soutien financier d’APIS-GENE dans le cadre du projet BEEFALIM 2020.
Références
- Adjorlolo, L., Obese, F. Y., & Tecku, P. (2019). Blood metabolite concentration, milk yield, resumption of ovarian activity and conception in grazing dual purpose cows supplemented with concentrate during the post‐partum period. Veterinary Medicine and Science, 5(2), 103‑111. doi:10.1002/vms3.148
- Alemu, A. W., Vyas, D., Manafiazar, G., Basarab, J. A., & Beauchemin, K. A. (2017). Enteric methane emissions from low– and high–residual feed intake beef heifers measured using GreenFeed and respiration chamber techniques1,2. Journal of Animal Science, 95(8), 3727‑3737. doi:10.2527/jas.2017.1501
- Amyes, N., & Morris, C. (2009). Responses to selection for pubertal traits in Angus cattle over 23 years. Proceedings of the New Zealand Society of Animal Production, 69, 201-205. https://www.nzsap.org/system/files/proceedings/2009/ab09048.pdf
- Atti, N. (1998). Effet du mode de conduite et de l’âge au sevrage de l’agneau sur les performances de production de la race laitière Sicilo-Sarde. Annales de l’INRAT, 71, 237‑249.
- Basarab, J. A., Colazo, M. G., Ambrose, D. J., Novak, S., McCartney, D., & Baron, V. S. (2011). Residual feed intake adjusted for backfat thickness and feeding frequency is independent of fertility in beef heifers. Canadian Journal of Animal Science, 91(4), 573‑584. doi:10.4141/cjas2011-010
- Berge, P. (1991). Long-term effects of feeding during calfhood on subsequent performance in beef cattle (a review). Livestock Production Science, 28(3), 179‑201. doi:10.1016/0301-6226(91)90142-D
- Berry, D. P., & Crowley, J. J. (2013). Cell Biology Symposium: genetics of feed efficiency in dairy and beef cattle. Journal of Animal Science, 91(4), 1594‑1613. doi:10.2527/jas.2012-5862
- Bidan, F., Le Mezec, P., Dimon, P., Salvetti, P., Lejard, A., Gelé, M., & Bareille, N. (2019). REPROSCOPE: the observatory of cattle reproductive performances in France. Proceedings ICAR Conference, 1‑13. https://www.icar.org/Documents/technical_series/ICAR-Technical-Series-no-24-Prague/Bidan.pdf
- Black, T. E., Bischoff, K. M., Mercadante, V. R. G., Marquezini, G. H. L., DiLorenzo, N., Chase, C. C., Jr., Coleman, S. W., Maddock, T. D., & Lamb, G. C. (2013). Relationships among performance, residual feed intake, and temperament assessed in growing beef heifers and subsequently as 3-year-old, lactating beef cows. Journal of Animal Science, 91(5), 2254‑2263. doi:10.2527/jas.2012-5242
- Blanc, F., Dumont, B., Brunschwig, G., Bocquier, F., & Agabriel, J. (2010). Robustesse, flexibilité, plasticité : des processus adaptatifs révélés dans les systèmes d’élevage extensifs de ruminants. INRA Productions Animales, 23(1), 65‑80. doi:10.20870/productions-animales.2010.23.1.3288
- Blanc, Fabienne, Bocquier, F., Agabriel, J., D’hour, P., & Chilliard, Y. (2006). Adaptive abilities of the females and sustainability of ruminant livestock systems. A review. Animal Research, 55(6), 489‑510. doi:10.1051/animres:2006040
- Brody, S. (1945). Bioenergetics on growth. Reinhold Publishing Corporation
- Butler, W. R. (2003). Energy balance relationships with follicular development, ovulation and fertility in postpartum dairy cows. Livestock Production Science, 83(2), 211‑218. doi:10.1016/S0301-6226(03)00112-X
- Callum, C., Ominski, K. H., Crow, G., Zvomuya, F., & Basarab, J. A. (2019). Relationship between residual feed intake classification as a heifer and lifetime productivity of beef cattle. Canadian Journal of Animal Science, 99(1), 191‑201. doi:10.1139/cjas-2018-0002
- Canépa, S., Lainé, A. L., Bluteau, A., Fagu, C., Flon, C., & Monniaux, D. (2008). Validation d’une méthode immunoenzymatique pour le dosage de la progestérone dans le plasma des ovins et des bovins. Cahier des Techniques de l’Inra, 64, 19‑30.
- Cantalapiedra-Hijar, G., Faverdin, P., Friggens, N. C., & Martin, P. (2021). Efficience Alimentaire : comment mieux la comprendre et en faire un élément de durabilité de l’élevage. INRAE Productions Animales, 33(4), 235‑248. doi:10.20870/productions-animales.2020.33.4.4594
- Cardoso, R. C., West, S. M., Maia, T. S., Alves, B. R. C., & Williams, G. L. (2020). Nutritional control of puberty in the bovine female: prenatal and early postnatal regulation of the neuroendocrine system. Domestic Animal Endocrinology, 73, 106434. doi:10.1016/j.domaniend.2020.106434
- Crowley, J. J., Evans, R. D., Mc Hugh, N., Kenny, D. A., McGee, M., Crews, D. H., & Berry, D. P. (2011). Genetic relationships between feed efficiency in growing males and beef cow performance. Journal of Animal Science, 89(11), 3372‑3381. doi:10.2527/jas.2011-3835
- De La Torre, A., Andueza, D., Renand, G., Baumont, R., Cantalapiedra-Hijar, G., & Nozière, P. (2019). Digestibility contributes to between-animal variation in feed efficiency in beef cows. Animal, 13(12), 2821‑2829. doi:10.1017/S1751731119001137
- De La Torre, A., Blanc, F., Egal, D., & Agabriel, J. (2010). Dynamique post-partum de l’évolution des réserves corporelles de la vache allaitante charolaise selon sa parité et l’alimentation hivernale. 17e Rencontres autour des Recherches sur les Ruminants, Paris. 17, 289-292. https://www.journees3r.fr/spip.php?article3016
- D’Hour, P., Petit, M., Pradel, P., & Garel, J. P. (1995). Évolution du poids et de la production laitière au pâturage de vaches allaitantes Salers et Limousines dans deux milieux. 2e Rencontres autour des Recherches sur les Ruminants, Paris. 2, 105‑108. https://www.journees3r.fr/spip.php?article1737
- Douhard, F., Tichit, M., Amer, P. R., & Friggens, N. C. (2014). Synergy between selection for production and longevity and the use of extended lactation: insights from a resource allocation model in a dairy goat herd. Journal of Animal Science, 92(11), 5251‑5266. doi:10.2527/jas.2014-7852
- Fitzsimmons, C., McGee, M., Keogh, K., Waters, S. M., & Kenny, D. A. (2017). Molecular physiology of feed efficiency in beef cattle. In: Scanes, C.G. & Hill, R.A.(Eds) Biology of domestic animals (p. 122-165). CRC Press. doi:10.1201/9781315152080-6
- Freetly, H. C., Nienaber, J. A., & Brown-Brandl, T. (2006). Partitioning of energy during lactation of primiparous beef cows. Journal of Animal Science, 84(8), 2157‑2162. doi:10.2527/jas.2005-534
- Fruscalso, V., Stumpf, M. T., McManus, C. M., & Fischer, V. (2013). Feeding restriction impairs milk yield and physicochemical properties rendering it less suitable for sale. Scientia Agricola, 70(4), 237‑241. doi:10.1590/S0103-90162013000400003
- Gabbi, A. M., McManus, C. M., Zanela, M. B., Stumpf, M. T., Barbosa, R. S., Fruscalso, V., Thaler Neto, A., Schmidt, F. A., & Fischer, V. (2016). Milk traits of lactating cows submitted to feed restriction. Tropical Animal Health and Production, 48(1), 37‑43. doi:10.1007/s11250-015-0916-2
- Giridhar, K., & Samireddypalle, A. (2015). Impact of Climate Change on Forage Availability for Livestock. In: Sejian,V., Gaughan, J., Baumgard, L. & Prasad, C. (Eds.) Climate Change Impact on Livestock: Adaptation and Mitigation (p. 97‑112). Springer eBooks. doi:10.1007/978-81-322-2265-1_7
- Grimard, B., Humblot, P., Ponter, A. A., Chastant, S., Constant, F., & Mialot, J. P. (2003). Efficacité des traitements de synchronisation des chaleurs chez les bovins. INRAE Productions Animales, 16(3), 211‑227. doi:10.20870/productions-animales.2003.16.3.3661
- Hafla, A. N., Carstens, G. E., Forbes, T. D. A., Tedeschi, L. O., Bailey, J. C., Walter, J. T., & Johnson, J. R. (2013). Relationships between postweaning residual feed intake in heifers and forage use, body composition, feeding behavior, physical activity, and heart rate of pregnant beef females. Journal of Animal Science, 91(11), 5353‑5365. doi:10.2527/jas.2013-6423
- Herd, R. M., Velazco, J. I., Arthur, P. F., & Hegarty, R. S. (2016). Proxies to adjust methane production rate of beef cattle when the quantity of feed consumed is unknown. Animal Production Science, 56(3), 231‑237. doi:10.1071/AN15477
- Kelly, A. K., Kenny, D. A., McGee, M., & Heslin, J. (2022). Morphological and physiological measures as predictors of age at puberty and conception in beef heifer genotypes. Applied Animal Science, 38(1), 22‑32. doi:10.15232/aas.2021-02205
- Koch, R. M., Swiger, L. A., Chambers, D., & Gregory, K. E. (1963). Efficiency of feed use in beef cattle. Journal of Animal Science, 22(2), 486‑494. doi:10.2527/jas1963.222486x
- Koonawootrittriron, S., Elzo, M. A., Tumwasorn, S., & Sintala, W. (2001). Lactation curves and prediction of daily and accumulated milk yields in a multibreed dairy herd in Thailand using all daily records. Thai Journal of Agricultural Science (Thailand). https://www.researchgate.net/publication/228914036
- Le Neindre, P., Petit, M., & Dubroeucq, H. (1975). Note sur l’estimation de la production laitière des vaches allaitantes au début de la lactation. Annales de zootechnie, 24(3), 559‑563. doi:10.1051/animres:19750315
- Lefebvre, R., Larroque, H., Barbey, S., Gallard, Y., Colleau, J. J., Lainé, A. L., Boichard, D., & Martin, P. (2021). Genome-wide association study for age at puberty and resumption of cyclicity in a crossbred dairy cattle population. Journal of Dairy Science, 104(5), 5794‑5804. doi:10.3168/jds.2020-18228
- Lepers, A., Aguerre, S., Promp, J., Taussat, S., Vinet, A., Martin, P., Philibert, A., Laramee, A., & Griffon, L. (2022). Paramètres génétiques de la vitesse de développement et du poids adulte asymptotique en bovins allaitants. 26e Rencontres autour des Recherches sur les Ruminants, Paris. 26, 203. https://journees3r.fr/spip.php?article5024
- Manafiazar, G., Zimmerman, S., & Basarab, J. A. (2016). Repeatability and variability of short-term spot measurement of methane and carbon dioxide emissions from beef cattle using GreenFeed emissions monitoring system. Canadian Journal of Animal Science, 97(1), 118‑126. doi:10.1139/cjas-2015-0190
- Martin, L. C., Brinks, J. S., Bourdon, R. M., & Cundiff, L. V. (1992). Genetic effects on beef heifer puberty and subsequent reproduction. Journal of Animal Science, 70(12), 4006‑4017. doi:10.2527/1992.70124006x
- Martin, P., Taussat, S., Vinet, A., Krauss, D., Maupetit, D., & Renand, G. (2019). Genetic parameters and genome-wide association study regarding feed efficiency and slaughter traits in Charolais cows. Journal of Animal Science, 97(9), 3684‑3698. doi:10.1093/jas/skz240
- Martin, P., Vinet, A., Allart, L., Launay, F., Dozias, D., Maupetit, D., & Renand, G. (2022). Influence of feed restriction and subsequent recovery on lactating Charolais cows. Livestock Science, 264, 105077. doi:10.1016/j.livsci.2022.105077
- Martin, P., Vinet, A., Launay, F., Dozias, D., Maupetit, D., & Renand, G. (2023). Short communication: Variability of response to feed restriction in lactating Charolais cows. Animal, 17(2), 100704. doi:10.1016/j.animal.2022.100704
- Mialon, M. M., Renand, G., Krauss, D., & Ménissier, F. (1998). Variabilité génétique de l’âge à la puberté chez la génisse charolaise et relation avec la vitesse de croissance. 5e Rencontres Rencontres autour des Recherches sur les Ruminants, 5, 119‑122. https://www.journees3r.fr/spip.php?article2305
- Minick Bormann, J., & Wilson, D. E. (2010). Calving day and age at first calving in Angus heifers. Journal of Animal Science, 88(6), 1947‑1956. doi:10.2527/jas.2009-2249
- Morris, C. A., Wilson, J. A., Bennett, G. L., Cullen, N. G., Hickey, S. M., & Hunter, J. C. (2000). Genetic parameters for growth, puberty, and beef cow reproductive traits in a puberty selection experiment. New Zealand Journal of Agricultural Research, 43(1), 83‑91. doi:10.1080/00288233.2000.9513411
- Mullins, Y., Keogh, K., Blackshields, G., Kenny, D. A., Kelly, A. K., & Waters, S. M. (2021). Transcriptome assisted label free proteomics of hepatic tissue in response to both dietary restriction and compensatory growth in cattle. Journal of Proteomics, 232, 104048. doi:10.1016/j.jprot.2020.104048
- Parsons, C. T., Dafoe, J. M., Wyffels, S. A., DelCurto, T., & Boss, D. L. (2021). The influence of residual feed intake and cow age on beef cattle performance, supplement intake, resource use, and grazing behavior on winter mixed-grass rangelands. Animals, 11(6), 1518. doi:10.3390/ani11061518
- Pereira, A. B. D., Utsumi, S. A., Dorich, C. D., & Brito, A. F. (2015). Integrating spot short-term measurements of carbon emissions and backward dietary energy partition calculations to estimate intake in lactating dairy cows fed ad libitum or restricted. Journal of Dairy Science, 98(12), 8913‑8925. doi:10.3168/jds.2015-9659
- Pereira, M. C. S., Dellaqua, J. V. T., Sousa, O. A., Santi, P. F., Felizari, L. D., Reis, B. Q., Pinto, A. C. J., Bertoldi, G. P., Silvestre, A. M., Watanabe, D. H. M., Estevam, D. D., Arrigoni, M. D. B., & Millen, D. D. (2020). Feedlot performance, feeding behavior, carcass and rumen morphometrics characteristics of Nellore cattle submitted to strategic diets prior the adaptation period. Livestock Science, 234, 103985. doi:10.1016/j.livsci.2020.103985
- Perry, G. A. (2016). Factors affecting puberty in replacement beef heifers. Theriogenology, 86(1), 373‑378. doi:10.1016/j.theriogenology.2016.04.051
- Petit, M., & Agabriel, J. (1993). État corporel des vaches allaitantes charolaises : signification, utilisation pratique et relations avec la reproduction. INRA Productions Animales, 6(5), 311‑318. doi:10.20870/productions-animales.1993.6.5.4212
- Phocas, F., & Sapa, J. (2004). Genetic parameters for growth, reproductive performance, calving ease and suckling performance in beef cattle heifers. Animal Science, 79(1), 41‑48. doi:10.1017/S1357729800054515
- Poppe, M., Bonekamp, G., van Pelt, M. L., & Mulder, H. A. (2021). Genetic analysis of resilience indicators based on milk yield records in different lactations and at different lactation stages. Journal of Dairy Science, 104(2), 1967‑1981. doi:10.3168/jds.2020-19245
- Puillet, L., Réale, D., & Friggens, N. C. (2016). Disentangling the relative roles of resource acquisition and allocation on animal feed efficiency: insights from a dairy cow model. Genetics Selection Evolution, 48(1), 72. doi:10.1186/s12711-016-0251-8
- Randel, R. D., & Welsh, T. H. (2013). Joint Alpharma-Beef Species Symposium: interactions of feed efficiency with beef heifer reproductive development. Journal of Animal Science, 91(3), 1323‑1328. doi:10.2527/jas.2012-5679
- Renand, G., Vinet, A., Decruyenaere, V., Maupetit, D., & Dozias, D. (2019). Methane and carbon dioxide emission of beef heifers in relation with growth and feed efficiency. Animals, 9(12), 1136. doi:10.3390/ani9121136
- Roche, J. R. (2007). Milk production responses to pre- and postcalving dry matter intake in grazing dairy cows. Livestock Science, 110(1), 12‑24. doi:10.1016/j.livsci.2006.08.016
- Rust, J. M. (2019). The impact of climate change on extensive and intensive livestock production systems. Animal Frontiers, 9(1), 20‑25. doi:10.1093/af/vfy028
- Sanz, A., Bernués, A., Villalba, D., Casasús, I., & Revilla, R. (2004). Influence of management and nutrition on postpartum interval in Brown Swiss and Pirenaica cows. Livestock Production Science, 86(1), 179‑191. doi:10.1016/S0301-6226(03)00165-9
- Schillo, K. K., Hall, J. B., & Hileman, S. M. (1992). Effects of nutrition and season on the onset of puberty in the beef heifer. Journal of Animal Science, 70(12), 3994‑4005. doi:10.2527/1992.70123994x
- Sepchat, B., D’Hour, P., & Agabriel, J. (2017). Production laitière des vaches allaitantes : caractérisation et étude des principaux facteurs de variation. INRA Productions Animales, 30(2), 139‑152. doi:10.20870/productions-animales.2017.30.2.2240
- Shaffer, K. S., Turk, P., Wagner, W. R., & Felton, E. E. D. (2011). Residual feed intake, body composition, and fertility in yearling beef heifers. Journal of Animal Science, 89(4), 1028‑1034. doi:10.2527/jas.2010-3322
- Tahir, M. S., Porto-Neto, L. R., Gondro, C., Shittu, O. B., Wockner, K., Tan, A. W. L., Smith, H. R., Gouveia, G. C., Kour, J., & Fortes, M. R. S. (2021). Meta-analysis of heifer traits identified reproductive pathways in Bos Indicus cattle. Genes, 12(5), 768. doi:10.3390/genes12050768
- Taussat, S., Vinet, A., Maupetit, D., Dozias, D., Chanteloup, L., Lainé, A.-L., Renand, G., & Martin, P. (2023). Effect of environmental conditions and genetics on sexual precocity of purebred Charolais heifers. Article soumis pour publication.
- Tucker, C. B., Lacy-Hulbert, S. J., & Webster, J. R. (2009). Effect of milking frequency and feeding level before and after dry off on dairy cattle behavior and udder characteristics. Journal of Dairy Science, 92(7), 3194‑3203. doi:10.3168/jds.2008-1930
- Villalba, D., Casasús, I., Sanz, A., Bernués, A., Estany, J., & Revilla, R. (2006). Stochastic simulation of mountain beef cattle systems. Agricultural Systems, 89(2), 414‑434. doi:10.1016/j.agsy.2005.10.005
- Villalba, D., Ripoll, G., Ruiz, R., & Bernués, A. (2010). Long-term stochastic simulation of mountain beef cattle herds under diverse management strategies. Agricultural Systems, 103(4), 210‑220. doi:10.1016/j.agsy.2010.01.003
- Yavas, Y., & Walton, J. S. (2000). Postpartum acyclicity in suckled beef cows: A review. Theriogenology, 54(1), 25‑55. doi:10.1016/S0093-691X(00)00323-X
Résumé
Dans un contexte économique et environnemental où l’élevage est de plus en plus questionné, la précocité, l’efficience alimentaire et la résilience sont devenus des points d’intérêt majeurs de par leur rôle potentiel pour améliorer l’efficience globale des élevages. Pour explorer ces caractères et les liens qui les unissent dans les systèmes allaitants, une expérimentation de grande envergure a été menée entre 2011 et 2021 sur 650 femelles charolaises. Cette expérimentation se découpait en trois périodes que les animaux effectuaient successivement : l’étude de la précocité sexuelle et de développement, l’étude de l’efficience alimentaire au travers de l’ingéré résiduel à deux ans, puis l’étude de la réponse à un challenge alimentaire (composé d’une phase de restriction et d’une phase de récupération) au cours de la lactation. Cet article regroupe l’ensemble des résultats obtenus. Si la précocité sexuelle définie par l’âge à la puberté est fortement liée à l’environnement (conduite, alimentation…) et notamment à la saison de naissance, l’héritabilité du caractère est néanmoins modérée (0,21 ± 0,08). Elle est également corrélée génétiquement avec un petit poids à la naissance et une forte croissance pré-sevrage. L’efficience alimentaire des animaux de deux ans est peu héritable (0,09 ± 0,08) mais corrélée favorablement avec la précocité. Face au challenge alimentaire, les vaches ont des réponses équilibrées entre perte de production laitière et mobilisation de réserves corporelles mais cette réponse moyenne de la race cache des disparités entre individus. Enfin, les génisses les plus efficientes présentaient des productions laitières amoindries une fois en lactation par rapport à leurs homologues inefficientes (jusqu’à − 0,7 kg de lait par jour en milieu de lactation soit environ 10 % de moins), ainsi que des retours en chaleurs plus long dans le cadre de la restriction alimentaire. Ces résultats promeuvent l’importance de considérer l’efficience sur le long terme en prenant en compte ses différentes composantes et de ne pas réaliser une sélection génétique sur une efficience alimentaire mesurée sur un moment court et unique pendant la phase de croissance.
Pièces jointes
Pas de document complémentaire pour cet articleStatistiques de l'article
 Vues: 3149
Vues: 3149
Téléchargements
 PDF: 635
PDF: 635
 XML: 97
XML: 97
Articles les plus lus par le même auteur ou la même autrice
- Gonzalo CANTALAPIEDRA-HIJAR, Philippe FAVERDIN, Nicolas C FRIGGENS, Pauline MARTIN, Efficience Alimentaire : comment mieux la comprendre et en faire un élément de durabilité de l’élevage , INRAE Productions Animales: Vol. 33 No 4 (2020)
- Aurélie MADRID, Renée DE CRÉMOUX, Luc DELABY, Hélène LARROQUE, Sandra NOVAK, Aurélie VINET, L’élevage de ruminants s’adaptera-t-il au changement climatique ? Impacts et leviers d’adaptation , INRAE Productions Animales: Vol. 38 No 2 (2025)
- Christian DUCROT, Maria Belén BARRIO, Alain BOISSY, François CHARRIER, Sergine EVEN, Pierre MORMÈDE, Sandrine PETIT, Marie-Hélène PINARD-VAN DER LAAN, François SCHELCHER, François CASABIANCA, Alain DUCOS, Gilles FOUCRAS, Raphaël GUATTEO, Jean-Louis PEYRAUD, Muriel VAYSSIER-TAUSSAT, Patrick VEYSSET, Nicolas C. FRIGGENS, Xavier FERNANDEZ, Améliorer conjointement la santé et le bien-être des animaux dans la transition des systèmes d’élevage vers la durabilité , INRAE Productions Animales: Vol. 37 No 3 (2024)
- Marie-Pierre SANCHEZ, Valérie WOLF, Cécile LAITHIER, Mohammed EL JABRI, Éric BEUVIER, Odile ROLET-RÉPÉCAUD, Nicolas GAUDILLIÈRE, Stéphanie MINÉRY, Yuliaxis RAMAYO-CALDAS, Thierry TRIBOUT, Alexis MICHENET, Mekki BOUSSAHA, Sébastien TAUSSAT, Sébastien FRITZ, Agnès DELACROIX-BUCHET, Philippe GROSPERRIN, Mickaël BROCHARD, Didier BOICHARD, Analyse génétique de la « fromageabilité » du lait de vache prédite par spectrométrie dans le moyen infrarouge en race Montbéliarde , INRAE Productions Animales: Vol. 32 No 3 (2019)
- Gonzalo CANTALAPIEDRA-HIJAR, Cécile MARTIN, Donato ANDUEZA, Milka POPOVA, Diego MORGAVI, Abimael ORTIZ-CHURA, Benoit GRAULET, Isabelle CASSAR-MALEK, Muriel BONNET, Anne DE LA TORRE CAPITAN, Gilles RENAND, Sébastien TAUSSAT, Isabelle ORTIGUES-MARTY, Pierre NOZIÈRE, Mécanismes digestifs et métaboliques associés aux différences inter-individuelles de l’efficience alimentaire chez le bovin allaitant , INRAE Productions Animales: Vol. 36 No 3 (2023): Dossier « Efficience alimentaire des bovins allaitants » & Articles hors dossier
- Florence GONDRET, Donato ANDUEZA, Mohamed Habibou ASSOUMA, Valérie BERTHELOT, Didier BOICHARD, Maguy EUGÈNE, Solène FRESCO, Amandine LURETTE, Cécile MARTIN, Pauline MARTIN, Diego P. MORGAVI, Rafael MUÑOZ-TAMAYO, Milka POPOVA, Simon ROQUES, Flavie TORTEREAU, Xavier FERNANDEZ, Réduction des émissions de méthane entérique chez les ruminants : enjeux, solutions et perspectives à l’échelle de l’animal et des systèmes d’élevage , INRAE Productions Animales: Vol. 39 No 1 (2026)
- Sébastien TAUSSAT, Clément FOSSAERT, Gonzalo CANTALAPIEDRA-HIJAR, Laurent GRIFFON, Pauline MARTIN, Gilles RENAND, Paramètres génétiques de l’efficience alimentaire et faisabilité d’une sélection en population bovine Charolaise , INRAE Productions Animales: Vol. 36 No 3 (2023): Dossier « Efficience alimentaire des bovins allaitants » & Articles hors dossier
- Sébastien TAUSSAT, Gonzalo CANTALAPIEDRA-HIJAR, Jacques AGABRIEL, Gilles RENAND, Sylvie BROUARD, Laurent GRIFFON, Pauline MARTIN, Caractériser les déterminants physiologiques et génétiques de l’efficience alimentaire des bovins allaitants : le programme BEEFALIM 2020 , INRAE Productions Animales: Vol. 36 No 3 (2023): Dossier « Efficience alimentaire des bovins allaitants » & Articles hors dossier
- Didier BOICHARD, Pascal CROISEAU, Florence PHOCAS, Romain SAINTILAN, François GUILLAUME, Sébastien TAUSSAT, Chris HOZÉ, Anne BARBAT, Aurélia BAUR, Hélène LECLERC, Carine CAPEL, Mathieu DIRIBARNE, Christine BERTRAND, Philippe BOULESTEIX, Mekki BOUSSAHA, Aurélien CAPITAN, Iola CROUÉ, Marine BARBAT, Romain DASSONNEVILLE, André EGGEN, Clémentine ESCOUFLAIRE, Armelle GOVIGNON, Cécile GROHS, Manon GUILLERM, Amandine LAUNAY, Marc LAVAL, Rachel LEFEBVRE, Laure-Hélène MAUGAN, Alexis MICHENET, Stéphanie MINERY, Marie-Pierre SANCHEZ, Aurélie VINET, Sophie MATTALIA, Eric VENOT, Clotilde PATRY, Thierry TRIBOUT, Vincent DUCROCQ, Sébastien FRITZ, Bilan de la sélection génomique bovine en France , INRAE Productions Animales: Vol. 39 No 2 (2026)
- Henri LAGARDE, Pauline MARTIN, Philippe BARRE, René BAUMONT, Mickaël BROCHARD, Philippine COEUGNET, Pascal CROISEAU, Luc DELABY, Anne DE LA TORRE, Nicolas C. FRIGGENS, Raphael GUATTEO, Frédéric JOLY, Hélène LECLERC, Marie-Angelina MAGNE, Sandra NOVAK, Christophe PERROT, Laurent SCHIBLER, Patrick VEYSSET, Aurélie VINET, Didier BOICHARD, Enjeux et orientations de la sélection génétique bovine en France d’ici 2040 : propositions issues d’une réflexion collective , INRAE Productions Animales: Vol. 39 No 2 (2026)