

Influence of pastoral management on the conservation of mountain pasture biodiversity (Full text available in English)
Today, mountain pastures, which provide an essential source of high-quality forage for livestock, are becoming increasingly fragile, along with their associated biodiversity and ecosystem services. Climate change and socio-economic changes are disrupting a balance that has existed for millennia. How can the management of these unique and enduring ecosystems be adapted in order to preserve them?
Introduction
Mountain meadows and pastures are one of the most diverse habitats in the world in terms of plant species richness (Dengler et al., 2014). In restricted zones (i.e. at plot-scale), their vascular plant diversity can even exceed that of tropical rainforests, generally considered to be the most diverse ecosystems on the planet. For example, at a scale of 1 m2, the world record of 89 plant species is held by the dry mountain grasslands of Central Argentina (Cantero et al., 1999). Furthermore, in plots of 16 and 49 m2, the highest vascular plant species richness (105 and 131 species respectively) was observed in semi-natural grassland situated in the Czech part of the White Carpathians (Wilson et al., 2012). Although the total number of species present in all European mountain meadows and pastures has yet to be quantified, some data are available for specific mountain ranges. For example, more than 75% of the 4,485 plant species recorded in the Alps are associated with grassland habitats situated from the hilly to the alpine belts (Aeschimann et al., 2013).
At global scale, the number of plant species in mountain regions generally increases with surface area, productivity and topographical variation (Rahbek et al., 2019). Several ecological factors help to explain the enormous biodiversity of these mountain zones: i) the ‘insular nature’ of mountains, which over time has favoured greater speciation compared with adjacent lowland zones, also favours a high number of endemic species (Körner & Spehn, 2024); ii) the high topographical variation (altitude, slope, orientation, roughness...), which has a direct or indirect influence on microclimatic factors (solar radiation, temperature, duration of snow cover, length of growing season...) and pedological factors (bedrock alteration, soil type, humidity, nutrient levels, accumulation of organic matter...); iii) the coexistence of natural grasslands above the treeline and semi-natural grasslands below the treeline, maintained by traditional farming practices such as livestock grazing or haymaking, which have characterised all European temperate grasslands since Neolithic times (Hejcman et al., 2013). Furthermore, the microtopographic variation is accompanied by a large variation in management practices, which amplifies the coexistence of different species within the same zone.
In Europe, the high biodiversity characterising mountain grassland habitats is greatest in semi-natural grasslands created from ancient woodland by human activity, which constitute an essential part of the European cultural landscape. Of these, dry, calcareous semi-natural grasslands (included in the Natura 2000 habitat of Community interest 6210*) are considered one of the most species-rich communities because they host several rare and endangered plant species, various bryophytes and lichens, and are of great importance for the conservation of the invertebrates associated with them, especially butterflies. In Switzerland, for example, they cover only 2.3% of the total grassland area but contain 13.1% of the Swiss flora and over half of all Swiss butterfly species, 170 of which depend entirely on this type of habitat (Masé, 2005).
When environmental conditions become more difficult for plants to survive, for example in nutrient-poor or very arid, stony environments, a reduction in the number of species can be observed as some species that are well-adapted to these difficult conditions become dominant at the expense of others. For example, in alpine pastures, very low and very high soil nutrient levels are generally associated with the lowest plant diversity; low nutrient levels encourage certain nutrient-poor plant species to become dominant within the plant community, while high nutrient levels promote the dominance of certain nitrophilous plant species (Figure 1). Likewise, at higher altitudes in alpine zones, abiotic environmental factors such as climatic parameters (duration of snow cover, for example) play a particularly important role in determining the assemblage of plant communities and have a significant influence on biodiversity (Marini et al., 2007). In mountain ecosystems, a general reduction in plant diversity can be seen with increasing altitude due to differences in temperature, seasonal growth, soil nutrient levels, deposition and mineralisation rates (Körner, 2021). If the ecological gradient studied is sufficiently large, the relationship between soil nutrient levels and plant species richness follows an asymmetric distribution of a hump-shaped curve which declines steeply towards the upper extremity of the gradient (Figure 1).
Figure 1: Grassland soil nutrient level and plant diversity
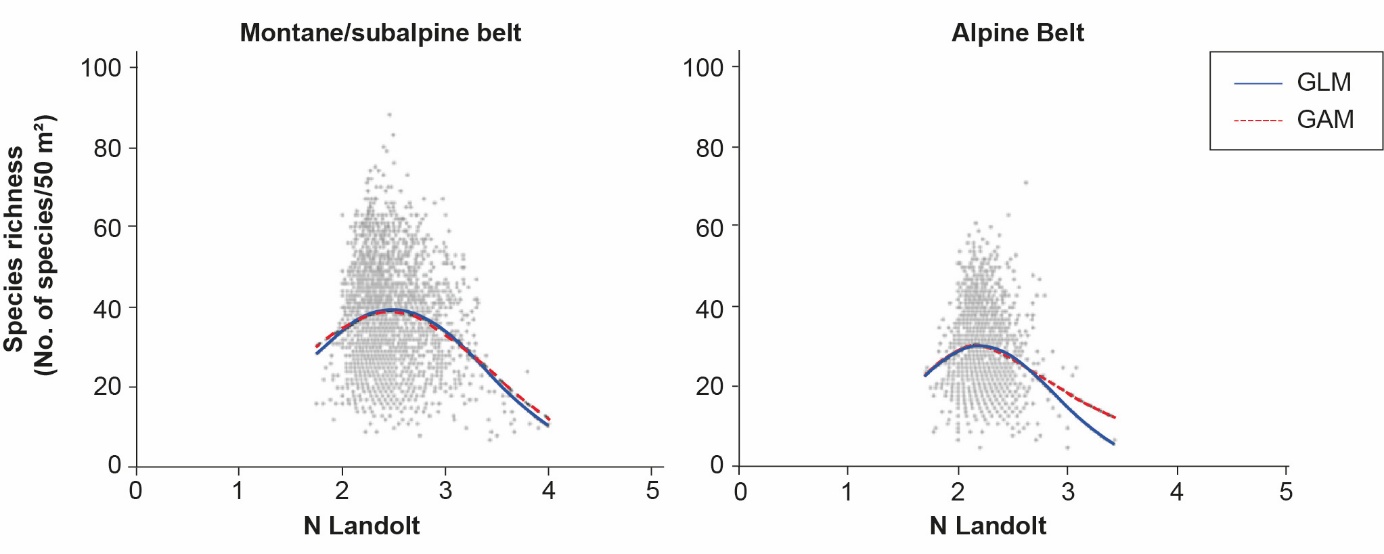
At landscape scale, the interaction between land use and site-specific environmental conditions is reflected in a diverse range of plant communities which contribute to a landscape characterised by high diversity (Gazol et al., 2012). European mountain meadows and pastures support a multitude of habitats and types of vegetation. For example, in the French Massif Central and the Western Italian Alps (Piedmont region), 135 and over 90 grassland types have been identified respectively, variably distributed according to altitude, soil nutrient level and management intensity (Cavallero et al., 2007; Le Hénaff et al., 2021).
1. Impact of livestock on plant diversity
Pastoral activities have fundamentally shaped mountain habitats. Early farmers cut down trees to keep warm and to make cheese, opening up the ancient wooded landscape, and continued to change it, for example by removing stones and creating terraces. Livestock have also played their part in modifying these habitats. Livestock grazing influences the environment directly and indirectly through selective defoliation, trampling, nutrient redistribution and seed transport. In this way, herbivores modify competition for light and soil nutrients between species within the plant community, thus shaping its botanic composition (Gaujour et al., 2012).
1.1. Selective defoliation
Essentially, livestock remove plant biomass through grazing. Thus plant species with a high capacity for regeneration and capable of withstanding regular defoliation have a competitive advantage over more vulnerable plants. Young shrubs and trees in particular are less adapted to grazing and have thus been progressively eliminated from meadows and pastures over the centuries. However, livestock do not graze uniformly (Bayle et al., 2019). Some patches are grazed intensively, creating ideal conditions for fast-growing species that require large amounts of light, for example. Other less grazed areas provide a refuge for slower growing, shade-adapted species. Insects and fauna also benefit from these uneven grazing patterns. For example, in the Alps, the presence of certain umbrella species indicating high plant and animal biodiversity, such as the black grouse, is closely linked to a highly heterogeneous mosaic of grassland and shrubby habitats (Braunisch et al., 2016).
This preference for one specific area of grassland and avoidance of another is not random but determined by the feeding preferences of different types of livestock (Wood, 1987). Livestock can be classed in three main categories: grazers, mixed feeders and browsers. Sheep and cattle are generally considered to be grazers because their diet consists mainly of grasses with less than 25% of herbaceous dicotyledons and woody plants, including the leaves and branches of shrubs. Cattle, however, use their tongues to tear forage, which makes them less selective than sheep and horses (Crofts, 1999), whose narrower mouths and more flexible lips enable them to graze closer to the ground. In contrast, thanks to the shape of their incisor arcade and their mobile lips, goats can generally include a higher percentage of woody species in their diet. As such, they are considered to be mixed feeders when dicotyledons comprise 25 to 75% of their diet, and browsers when dicotyledons comprise more than 75% of their diet (Iussig et al., 2015).
Selective defoliation has a direct impact on grassland vegetation, shaping both the growth form of different species and the botanical composition (Díaz et al., 2007). Intensive grazing modifies the growth form of different plant species in the short and medium term, leading to a general reduction in height and total biomass produced and an increase in prostrate growth forms. In the medium and long term, the botanical composition is also affected as the proportion of species resistant to defoliation (productive grasses, for example) increases and fast-growing plant species with a high potential for regeneration outcompete more vulnerable species (Díaz et al., 2007). Rosette-forming or stoloniferous plants (unlike tussock grasses) are normally spared because they are too short to be reached by grazers’ mouths (Díaz et al., 2007). Furthermore, they benefit from the elimination of taller plants, which gives them more light and reduces competition for nutrients. Some plants have developed specific defence mechanisms to avoid defoliation, such as toxic secondary compounds, unpalatable structures (such as tough, prickly leaves or thorns) or very low nutrient levels in their above-ground biomass (Pauler et al., 2020a). In summary, livestock have created a broad range of ecological niches occupied by a multitude of plant species exhibiting highly variable adaptation mechanisms in response to grazing. However, if the intensity and frequency of defoliation is too high in the long term, the heterogeneity of the conditions, and thus plant diversity, diminishes. The underlying mechanism is that a few very well-adapted plants overgrow all other species. They become exceptionally dominant within the plant community, thereby reducing overall plant diversity. In effect, according to the intermediate disturbance hypothesis, species richness is the highest at moderate levels of grazing intensity, since it peaks at intermediate values of disturbance due to the coexistence of several species in overlapping ecological niches (Grime, 1973).
Furthermore, over the course of the last century, humans have greatly modified the characteristics of cattle, thereby, unintentionally disrupting the delicate balance between the preferences of different breeds for and their avoidance of certain plant species. Modern livestock breeding has favoured high milk and meat production, and this increase in productivity is accompanied by an increased demand for nutrients. To meet this demand, modern breeds select positively for plant species with a high nutritional value, such as broad-leaved grasses and legumes. In contrast, they clearly avoid less palatable and less digestible plants, as these animals are unable to meet their genetically defined demand for meat or milk production by selecting forage plants with low nutritional value without receiving supplementary feed concentrates (Berry et al., 2002; Pauler et al., 2020a). In addition, modern breeds make more effort than traditional breeds to avoid plant species that protect themselves from herbivores. For example, thorny species such as thistles, or toxic plants, are more often selected by less productive Highland cattle than by higher-yielding modern breeds (Pauler et al., 2020a). This modifies the delicate balance between preference and avoidance: the competitive advantage of less palatable plant species increases and they become more and more dominant, overgrowing other plant species. For example, in high-altitude grasslands, the less palatable grass species Nardus stricta L. is invading ancient biodiverse habitats because modern breeds tend to avoid it. This phenomenon is accompanied by a decline in the biodiversity and forage quality of these grasslands. By improving livestock productivity, modern farming has inadvertently increased their foraging selectivity and grassland uniformity while at the same time reducing grassland biodiversity (Pauler et al., 2019).
1.2 Trampling
Trampling by livestock has a significant, direct influence on the botanical composition of grasslands. Plant species vulnerable to trampling can be destroyed by the pressure of the animals’ hooves. However, some species have evolved specific physiological adaptations in response to trampling, such as rosettes, stolons or rhizomes (Díaz et al., 2007) which enable them to conserve nutrients safely underground and regenerate rapidly after trampling. These trampling specialists benefit from grazing activity because their competitors are weakened or destroyed in zones frequently visited by livestock, while other seldom visited areas of alpine pasture offer a refuge for species less adapted to trampling. For example, less abundant subordinate species – a key component of plant diversity in grasslands – benefit from moderate trampling because the gaps created by cattle reduce root competition from dominant species (Mariotte et al., 2013). In this way, moderate livestock trampling can increase the heterogeneity and thus the biodiversity of alpine pastures, not only for plants, but for other organisms as well. Amphibians, for example, can benefit from water-filled hoof prints, soil compaction, which reduces evapotranspiration, or the elimination of senescent vegetation, creating a supplementary habitat and foraging opportunities (Howell et al., 2019). On the other hand, if grazing intensity is too high, they can suffer from increased levels of nitrate and dissolved oxygen, and sediment loads (Howell et al., 2019).
However, the beneficial effect of trampling applies only to well-managed grasslands where grazing intensity is site-adapted. In mountain regions, the rugged topography often causes livestock to gather in more favourable zones, such as flat resting areas, zones around barns, points of attraction (water troughs, mineral supplements) and tracks following the contours (Probo et al., 2014). If trampling pressure is not evenly spread across the pastures, species richness in plant communities in the most trampled zones diminishes. As a result, these communities become dominated by a small number of species which tolerate the repeated passage of livestock, such as Taraxacum officinale aggr., Alchemilla vulgaris aggr., Plantago major L., Poa supina Schrad. and Poa trivialis L. (Supek et al., 2014). If grazing and trampling pressure increases further, especially in wet conditions, the vegetation can be destroyed. Ground laid bare in this way can be rapidly colonised by species adapted to trampling, such as Rumex alpinus L, which spreads across bare ground more rapidly than other species. These species not only diminish the forage quality of grassland; they also reduce biodiversity by outcompeting species less resistant to trampling and thus dominating the plant community.
As we have already mentioned, modern livestock breeding has modified livestock traits, which in turn has led to a more negative impact on alpine pastures: cattle bred for production are heavier than traditional breeds, thereby increasing the body weight pressure to the ground. Furthermore, their hooves are comparatively small relative to their body weight. The pressure per square centimetre exerted on the ground is thus far greater than with traditional breeds. Pauler et al. (2020b) stated that the pressure exerted by the hooves of a highly productive Angus × Holstein crossbreed is more than one third higher than that exerted by the hooves of the robust, less productive Highland cattle (Pauler et al., 2020b). Higher trampling pressure exacerbates the negative effects of trampling on biodiversity (Pauler et al., 2019). Moreover, highly productive breeds of cattle use the available space less evenly than traditional breeds: the more productive the cattle, the more time they spend in flat, nutrient-rich pastures, while avoiding steep, nutrient-poor areas. In contrast, robust breeds such as Highland cattle also explore steep, stony areas where forage quality is low (Pauler et al., 2020b). By clustering in restricted areas, modern breeds often overgraze these more attractive zones, leading to soil compaction and destruction of the vegetation. Thus, modern livestock breeding has aggravated the negative impact of trampling and increased the risk of erosion at local level in mountain zones.
1.3 Redistribution of seeds and nutrients
Apart from slope, other factors determine the distribution patterns of cattle in mountain pastures, such as quality and quantity of forage or distance from infrastructures and watering points (Probo et al., 2014; Homburger et al., 2015). Given that grazing cattle do not use the available space evenly, they drive a redistribution of nutrients within the pastures. In effect, nutrients in alpine pastures are transported mainly through animal excretions; nutrients absorbed during foraging are returned through urine and dung (Schnyder et al., 2010). The spatial distribution of dung and urine is not uniform; excretions are mainly deposited in flat, localised resting areas, which become nutrient-enriched over time, while steeper pastures are characterised by nutrient depletion (Svensk et al., 2023). Nutrient availability (especially of nitrogen) strongly affects the development of plant species as it is one of the most important limiting factors for plant species and communities in mountain environments, affecting both grassland productivity and the composition of plant communities. By creating zones with contrasting nutrient levels, grazing cattle can increase the heterogeneity and biodiversity of grassland at landscape level, on a gradient ranging from nutrient-poor to nutrient-rich plant communities. Thus nutrient-rich zones dominated by tall, nitrophilous generalists can be found alongside nutrient-poor zones harbouring small, undemanding specialists.
Furthermore, livestock act as vectors for seed dispersal both within and between pastures, mainly by endozoochory, i.e. ingesting seeds and excreting them in faeces, and epizoochory, i.e. transporting seeds externally. This latter method is particularly important for certain types of livestock such as sheep. These zoochoric plant species are particularly impacted by a reduction in stocking levels. In addition, in pastures that are no longer grazed, zoochoric plant species that rely on dispersal by animals lose their connection to other populations. These isolated populations have a weaker survival capacity. Seed dispersal by livestock is thus an important and often neglected aspect of maintaining plant diversity. It has been shown that long-haired Highland cattle increase biodiversity by transporting the seeds of epizoochoric plant species (Pauler et al., 2019).
2. Impact of grazing management on plant diversity
2.1. Intensification and extensification process
Species-rich pastures created by livestock are not self-sustaining; they depend on continuous grazing as part of a site-adapted management strategy. However, two opposing trends have emerged in European mountain pastures in recent decades, both of which have a harmful effect on plant diversity: intensification of favourable zones and extensification of the most marginal zones (Tasser & Tappeiner, 2002).
Pastures located in favourable conditions, such as flat zones with high-quality forage, good infrastructure and proximity to roads and farm buildings, have generally been managed more intensively. This is reflected in higher stocking rates and/or an extended grazing season, the selection of more productive breeds, the replacement of small ruminants with cattle and the use of fertilisers and machines. These measures have led to a homogenisation of grassland communities. The ecological niches occupied by plant and animal species adapted to extensive systems have been reduced along with the overall biodiversity of these zones. In intensively managed pastures, there has been a shift towards a few generalist plant species which are best equipped to exploit these conditions. If management intensity increases further and pastures become overgrazed, high trampling pressure leads not only to biodiversity loss but also to compaction and discontinuity of the soil cover. This increases the risk of soil instability and erosion. Furthermore, a nutrient surplus concentrated in favourable areas leads to nutrient leaching, thereby reducing water quality downstream.
The reverse has occurred in less favourable pastures: often, farmers have not been able to make a profit on steep, rocky and remote areas with poor-quality forage. Climate change has accelerated this process, with a reduction in glacial water flow and an increase in summer droughts leading to a lack of water for animals in many regions. Due to their low profitability, these pastures have often been managed extensively, in other words grazed with very low stocking rates, or even abandoned altogether. The abandonment of pastures favours a small number of dominant plant species, resulting in a sharp decline in diversity (Mariotte et al., 2012). The reduction in grazing has also fundamentally changed the factors determining the composition of grassland plants: the succession of woody species starts after abandonment when livestock are no longer continually defoliating grassland plants and trampling on young saplings. Shrub encroachment and reforestation are accompanied by the loss of several ecosystem services provided by alpine pastures, such as traditional meat and cheese production or the creation of an attractive landscape that is of value to tourism and the economy (Soliva et al., 2010). However, this succession from open, species-rich mountain grasslands to scrub and forest is caused not only by a reduction in livestock but by a change in the type of livestock. In Switzerland for example, the number of goats has declined by 80% in the last 150 years, while the number of cattle has increased by 50% in the same period (Pauler et al., 2022). With grazers having replaced browsers, there are fewer animals to debark woody plants and so reduce their cover.
Figure 2: Woody species cover and plant diversity.
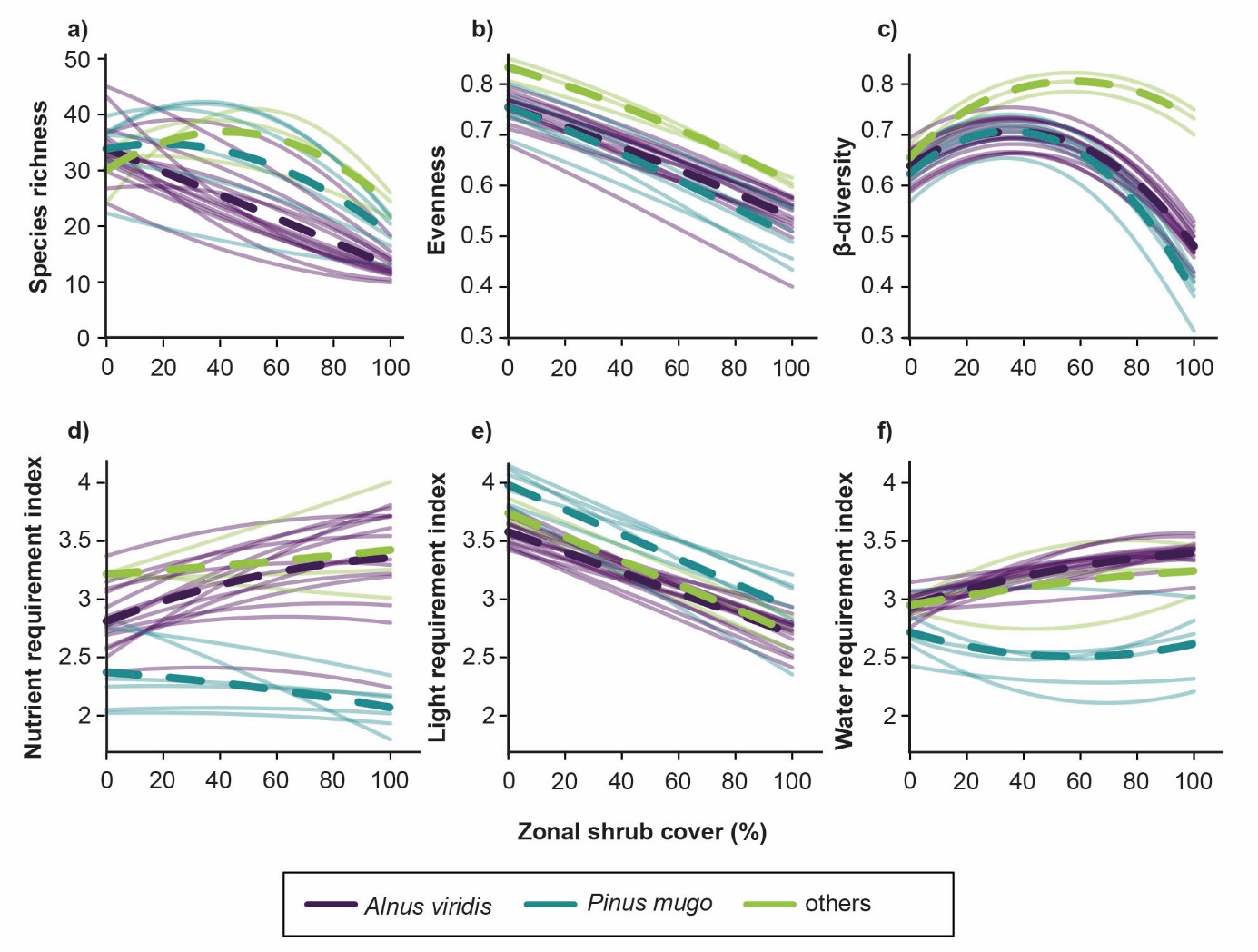
Biodiversity benefits from extensification provided there is a mosaic of open grassland and isolated shrubs and trees: plant species richness reaches its peak when shrub cover is low to moderate (Figure 2). However, if grazing is further reduced and woody species become dominant, biodiversity diminishes due to the much greater uniformity of the woody vegetation. This applies equally to dwarf shrubs such as Rhododendron ferrugineum L. (Pornaro et al., 2017) and tall ones such as Pinus mugo Turra. There is one exception to this hump-shaped relationship between shrub cover and biodiversity: in pastures colonised by green alder (Alnus viridis Chaix), even very low shrub cover results in a reduction in biodiversity, which diminishes linearly with each additional shrub (Figure 2). Unfortunately, green alder spreads more rapidly than any other shrub in the European Alps and mountain ranges of Eastern Europe due to its ability to fix atmospheric nitrogen (Pauler et al., 2022). The surplus nitrogen creates a eutrophic understory where a few nitrophilic plant species outcompete all others. In addition, nitrogen enrichment over-fertilises neighbouring pastures and downstream rivers and streams – thereby reducing biodiversity in the surrounding area – and is emitted to atmosphere as nitrous oxide, which is an important greenhouse gas.
The use of adapted livestock management to control green alder can make a very effective contribution to maintaining plant biodiversity (Svensk et al., 2022). The impact of shrub encroachment on other taxa is more complex. For example, the abandonment of grazing land leads to an overall increase in avian diversity. However, most species that colonise abandoned pastures are common and widespread, whereas endangered grassland bird species decline in number due to habitat loss (Laiolo et al., 2004).
2.2 Grazing systems
The way livestock is managed has a fundamental impact on the botanical composition of grasslands. Historically, the mountains of Europe were home to a large number of small, family-owned herds and grazing systems were typically based on herding and shepherding (MacDonald et al., 2000). These systems often allowed the controlled management of feed resources, with the animals led daily to grazing areas where forage species were often at the optimal phenological stage. These herding practices also made it possible to exploit the climatic altitudinal gradient and vertical transhumance of livestock within these mountain ecosystems (Meuret & Provenza, 2015). However, socio-economic and structural changes have profoundly modified these pastoral systems in recent decades. In mountain areas, the number of farms and the average number of workers per farm has declined sharply while the average number of animals per herd has risen to maintain an adequate level of profitability. Consequently, to reduce labour requirements, continuous grazing systems have often replaced traditional herding practices in several mountain areas such as the Italian Alps and Apennines (Probo et al., 2014). In continuous grazing systems, livestock are released and permitted to roam free on vast areas of grassland. The practice of livestock roaming free on mountain terrain has led to a more selective and spatially heterogeneous use of grassland than in the past when shepherding limited the animals’ natural preference for gentle topographies with more productive plants of higher nutritional value. Indeed, herders encouraged livestock to graze on steep terrain (Meuret & Provenza, 2015). Consequently, continuous grazing systems have exacerbated the processes of under- and overgrazing, with negative impacts on the conservation of biodiversity (Probo et al., 2014). Rotational grazing systems are an alternative management practice which can mitigate this problem. These systems involve moving livestock systematically from one fenced paddock to another while leaving other areas ungrazed. To reduce selective grazing by livestock, grazing duration and stocking rates in these zones should be balanced with forage availability, as this enables better management of forage resources and has a positive effect on forage quality (MacDonald et al., 2000). Perotti et al. (2018) showed that the implementation of a rotational grazing system in a high-altitude alpine pasture over a five-year period had a beneficial effect on biodiversity conservation by promoting seed transfer and increasing connectivity amongst different plant communities.
2.3. Stocking rates and frequency, timing and duration of grazing
One of the most important aspects of grazing management in terms of its effects on changes in plant diversity in mountain pastures is the stocking rate, in other words the number of livestock units grazing a specific zone during a given period (Dumont et al., 2009). It is acknowledged that a stocking rate that balances animal needs and vegetation carrying capacity would maximize both animal and pasture health, also in terms of plant species richness (Pittarello et al., 2021). Vegetation carrying capacity can be estimated by means of different techniques, such as the execution of phyto-pastoral vegetation surveys aiming at assessing the botanical composition, its associated forage pastoral value and terrain characteristics according to the procedures described in Cavallero et al. (2007). Often, however, the stocking rate is imbalanced and does not match vegetation carrying capacity. Stocking rates that are too high or too low in relation to vegetation carrying capacity have a negative impact on the botanical composition of the grassland, respectively leading to the development of nitrophilic plant communities or shrub encroachment. More specifically, imbalanced, too high stocking rates generally result in an increase in competitive and ruderal species (especially annuals), a reduction in plant height (including rosette structures), early flowering and seed-dispersing species. In contrast, imbalanced, low stocking rates can favour stress-tolerant grasses and herbaceous plants (Gaujour et al., 2012) as well as shrub encroachment (Probo et al., 2014).
The stocking rate impacts on vegetation depend not only on the number of animals per unit of surface and time and their relation with vegetation carrying capacity, but also on the frequency with which the grazing cycle is repeated over time. In fact, in numerous situations, especially in rotational grazing systems, foraging activities can be repeated within the same grazing season. In this case, care must be taken to ensure that the grass has had sufficient time to recover and produce enough biomass to be grazed again. Grazing frequency – presumably linked to grazing intensity – has a significant effect on the plant structure and diversity of each distinct parcel of vegetation. Initially, livestock grazing can bring about local changes in the composition of the plant community which may become permanent if the same grazing regime is repeated in the long term. Furthermore, frequent grazing favours the presence of species with high resistance to defoliation, as shown by the grazing and trampling indices proposed by Briemle et al. (2002). In contrast, low grazing frequency is often associated with higher plant diversity (Ravetto Enri et al., 2020a), which supports the intermediate disturbance hypothesis. Homburger et al. (2015) stated that a strict pasture rotation involving short periods of grazing is essential to influence livestock activity and hence effects on ecosystem processes and vegetation dynamics. Thus, precisely managed grazing has a positive effect on the provision of ecosystem services and consequently on biodiversity.
INRAE (French National Research Institute for Agriculture, Food and Environment) has conducted several studies in the Massif Central to evaluate the effectiveness of innovative grazing techniques in improving trophic availability for insect pollinators (Farruggia et al., 2012; Ravetto Enri et al., 2017). According to these studies, excluding a plot from a rotational grazing system, equivalent to a short period of rest during the main flowering season for dicotyledons, significantly increases butterfly and bumblebee abundance. It should be noted that the measures were not shown to significantly reduce grassland or animal performance, underlining the possibility of integrating nature conservation and the needs of livestock in an innovative ‘biodiversity-friendly’ grazing system.
The typical herd management systems over European mountains are based on vertical transhumance and they are based on a progressive exploitation of the pastures at different altitudes, encompassing a first grazing event at lower altitudes at the end of spring/early summer, followed by a shift of livestock to the summer pastures located at the highest elevations, and a second grazing event on vegetation regrowth at the end of the summer grazing season/beginning of autumn at lower altitudes. This system ensures that low-altitude vegetation has a sufficiently long rest, which can be beneficial for the plants’ flowering cycle and for habitat and biodiversity conservation. On the Pyrenees, Ubach et al. (2023) found that butterfly species richness and abundance increased during summer when the herd moved towards the higher pastures.
Early grazing is generally recommended to control the growth of shrubs, weeds and poor-quality plants (Dörner, 2023), as it hampers completion of the reproductive cycle and subsequent propagation of poor-quality forage species by seed dispersal, while at the same time stimulating the growth of high-quality species (especially grazing-tolerant grasses) through tillering. However, the palatability of low-quality forage species diminishes as the growing season progresses, mainly due to the rising fibre content and falling protein levels and digestibility. Consequently, early grazing is recommended for the optimal management of marginal zones characterised by nutrient-poor vegetation.
The timing and length of the grazing season should also cope with climate change, which is highly impacting on mountain pasture vegetation and the related pastoral management. Ravetto Enri et al. (2020), for instance, recommended adaptive exploitation dates for the management of Swiss species-rich grasslands instead of predefined fixed dates, in order to take into account the mean phenological stage of selected key species, to consider interannual weather fluctuations and the ongoing anticipation of vegetation phenology.
In addition, specific grazing regimes can be implemented to manage alpine pastures and their biodiversity. In particular, ‘temporary night camp areas’ with a high stocking rate can be introduced on undergrazed, shrub-encroached areas (Pittarello et al., 2016). Livestock can be attracted to these zones though the strategic placement of mineral-based (Probo et al., 2013) or molasses-based blocks (Svensk et al., 2022). The concentration of livestock activities (defoliation, trampling, transportation of seed and nutrients) in temporary night camp areas and around points of attraction can be particularly effective in reducing shrub cover and increasing herbaceous cover in the medium term (Probo et al., 2016). A study conducted in the western Alps found that the botanical composition changed significantly over a three-year period, with an increase in plant diversity in the temporary night camp areas (Pittarello et al., 2016). Specifically, this management regime was found to increase the cover of meso-eutrophic grassland and fringe and tall herb species, while markedly decreasing shrub and woodland cover.
Conclusions and outlook
As numerous examples in the Alps have shown, site-adapted grazing management which avoids over- and undergrazing is key to promoting the biodiversity of mountain pastures. One of the factors that has the greatest impact on the biodiversity of alpine pastures today is the implementation of direct payment schemes designed to govern farming practices. Direct payment schemes define rules for minimum stocking rates, grazing dates and specific grassland management regimes (for example, shrub clearance, weed control...). These policies vary from one country to another and in different regions of countries in the European Union in line with rural development plans. Often, however, they are not geared to the specific conditions of different mountain sites and merely define generic thresholds for individual parameters (stocking rates, for example) that must be met to access the direct payments. As they currently stand, direct payments cannot prevent shrub encroachment and the abandonment of unfavourable zones, and their implementation is controversial. This process is further exacerbated by growing pressure from large carnivores which are reclaiming mountain areas (Boitani & Linnell, 2015). On difficult terrain, protective measures such as extra fencing, night camp areas, permanent shepherds and livestock guardian dogs are costly and time-consuming. These measures are difficult, if not impossible to implement, especially in the steep, remote areas most at risk of abandonment. Although farmers often receive compensation for the loss of livestock killed by wolves or bears (Dalmasso et al., 2012), they seldom graze their animals in regions where predator levels are known to be high. Small ruminants, which would be the most efficient way to control shrub encroachment and prevent biodiverse pastures, are especially exposed to wolf attacks and thus their number is reduced most (Pauler et al., 2022).
Climate change also raises questions about the management of alpine pastures, and thus biodiversity. Rising temperatures and evaporation rates will further complicate the management of water and the maintenance of site-adapted grazing pressure in some areas. On the other hand, the availability of forage biomass will increase at higher altitudes where temperature has until now been a limiting factor (Guggenberger et al., 2021). Consequently, it will be necessary to increase grazing pressure to avoid undergrazing and its negative consequences for biodiversity. At the same time, more prolonged periods of drought will reduce the availability of fresh forage and force farmers to develop innovative adaptation strategies to fill the forage gap and avoid overgrazing during droughts.
There are various ways of responding to the multiple challenges facing mountain pastures and their biodiversity. Firstly, results-based policies can promote new site-adapted management strategies aimed at preserving the biodiversity of alpine pastures. In a recent study undertaken in the Apennines and the Alps, Napoleone et al. (2022) highlighted that grazing contracts at farm level and periodic field monitoring can have a positive influence on fine-scale grazing intensity. This would facilitate the transition from a rules-based approach to a results-based approach, in which the desired outcome could be the persistence of certain species typical of the habitats in question, for example. The large-scale implementation of these systems could allow direct control of the environmental impact of payments and raise the environmental awareness of farmers, encouraging them to gradually and voluntarily adopt measures to conserve biodiversity and the associated ecosystem services (Russi et al., 2016).
Secondly, one climate change adaptation strategy could be the development of silvopastoral systems where grassland is combined with tall-canopy trees which provide shade and so reduce the negative impacts of drought on herbaceous biomass and thermal stress on the animals. In addition, several tree species could supply an additional source of forage, as they are more resistant to periods of summer drought and their leaves provide good forage quality (Ravetto Enri et al., 2020b). Furthermore, silvopastoral systems characterised by a mosaic of trees, shrubs and grassland can create several microclimates which can support different plant habitats and communities, thereby increasing overall biodiversity at farm- and landscape-scale, as shown in the woodland pastures of the Jura (Buttler, 2014). In this case too, site-adapted management must provide long-term grazing systems designed not only to maintain the botanical composition of the herbage, but also to regenerate the forest component. Active grazing management of extensive areas that have suffered shrub encroachment in recent decades could help these shrub forests evolve to become silvopastoral systems characterised by a patchwork of grassland and tall-canopy trees. Active management of the forage resource provided by the understory could also minimise the risk of fires. To achieve this outcome, however, forest management policies and direct payment schemes for agriculture, which are currently at odds, would have to be harmonised.
Thirdly, the transfer of scientific knowledge to farmers is crucial in an age when traditional knowledge is rarely passed on orally. Practitioners can preserve the biodiversity of their mountain pastures only when knowledge about the long-term management of grassland is collected and made easily accessible to them. Swiss alpine pasture experts have created a website that explains how to control problematic weeds and shrubs (https://www.patura-alpina.ch/): the fact sheets and videos featuring farmers and experts suggest long-term measures that go beyond the use of herbicides.
And finally, new technologies can facilitate site-adapted management and conserve the biodiversity of mountain pastures. Airborne sensing systems already provide information about the condition of grassland, such as the amount of available biomass, but are not yet applicable to mountain zones with a heterogenous structure of vegetation. Geographic positioning systems and supplementary sensors on the animals allow farmers to track their animals’ whereabouts and see what conditions they are encountering in real time. Virtual fencing reduces the workload associated with installing fences on difficult terrain and allows livestock to be managed with greater precision (Probo et al., 2024a), for example, by excluding vulnerable, species-rich grazing zones on wet days, where trampling damage could be exacerbated. However, most of these new systems have been designed for interior or lowland applications and for highly productive livestock, and do not always work effectively in mountain conditions. Furthermore, mountain areas invariably have specific limitations, such as poor mobile phone coverage, which make it more difficult to introduce new technologies. Modern technologies are also associated with high costs, so it should be decided on a case-by-case basis whether they make sense in mountain systems.
Authors’ contributions
MP: conceptualisation, methodology, supervision, validation, drafting of the original and revised version, and correction; ML: conceptualisation, methodology, drafting of the original version; SRE: conceptualisation, methodology, drafting of the original and revised version, and correction; PM: conceptualisation, methodology, drafting of the original and revised version, and correction; CP: conceptualisation, methodology, drafting of the original and revised version, and correction.
Acknowledgments
The authors thank Carol Finch for the English language translation.
Notes
- 1. This article was presented at the 27e journées Rencontres autour des Recherches sur les Ruminants, 4-5 December 2024 in Paris (Probo et al., 2024b).
References
- Aeschimann, D., Rasolofo, N., & Theurillat, J.-P. (2013). Analyse de la flore des Alpes. 5 : Milieux et phytosociologie. Candollea, 68(1), 5-27. doi:10.15553/c2013v681a1
- Bayle, A., Carlson, B. Z., Thierion, V., Isenmann, M., & Choler, P. (2019). Improved mapping of mountain shrublands using the sentinel-2 red-edge band. Remote Sensing, 11(23), 2807. doi:10.3390/rs11232807
- Berry, N. R., Jewell, P. L., Sutter, F., Edwards, P. J., & Kreuzer, M. (2002). Selection, intake and excretion of nutrients by Scottish Highland suckler beef cows and calves, and Brown Swiss dairy cows in contrasting Alpine grazing systems. The Journal of Agricultural Science, 139(4), 437-453. doi:10.1017/S002185960200271X
- Boitani, L., & Linnell, J. D. (2015). Bringing large mammals back: large carnivores in Europe. In H. Pereira & L. Navarro (Eds.), Rewilding European Landscapes (pp. 67-84). Springer Open. https://doi.org/10.1007/978-3-319-12039-3
- Braunisch, V., Patthey, P., & Arlettaz, R. (2016). Where to combat shrub encroachment in alpine timberline ecosystems: Combining remotely-sensed vegetation information with species habitat modelling. PloS One, 11(10), e0164318. doi:10.1371/journal.pone.0164318
- Briemle, G., Nitsche, S., & Nitsche, L. (2002). Nutzungswertzahlen für Gefässpflanzen des Grünlandes. Schriftenreihe für Vegetationskunde, 38(2), 203-225.
- Buttler, A. (2014). Grasslands in silvopastoral mountain ecosystems. In P. Mariotte & P. Kardol (Eds.), Grassland biodiversity and conservation in a changing world (pp. 187-218). Nova Science Publishers.
- Cantero, J. J., Pärtel, M., & Zobel, M. (1999). Is species richness dependent on the neighbouring stands? An analysis of the community patterns in mountain grasslands of central Argentina. Oikos, 87(2), 346-354. doi:10.2307/3546750
- Cavallero, A., Aceto, P., Gorlier, A., Lombardi, G., Lonati, M., Martinasso, B., & Tagliatori, C. (2007). I tipi pastorali delle Alpi piemontesi: Vegetazione e gestione dei pascoli delle Alpi occidentali. Alberto Perdisa Editore.
- Crofts, A. (1999). The lowland grassland management handbook. English Nature & The Wildlife Trusts. https://publications.naturalengland.org.uk/publication/35034
- Dalmasso, S., Vesco, U., Orlando, L., Tropini, A., & Passalacqua, C. (2012). An integrated program to prevent, mitigate and compensate wolf (Canis lupus) damage in Piedmont region (northern Italy). Hystrix, the Italian Journal of Mammalogy, 23(1), 54-61. https://doi.org/10.4404/hystrix-23.1-4560
- Dengler, J., Janišová, M., Török, P., & Wellstein, C. (2014). Biodiversity of Palaearctic grasslands: a synthesis. Agriculture, Ecosystems & Environment, 182, 1-14. https://doi.org/10.1016/j.agee.2013.12.015
- Díaz, S., Lavorel, S., McIntyre, S., Falczuk, V., Casanoves, F., Milchunas, D. G., Skarpe, C., Rusch, G., Sternberg, M., Noy‐Meir, I., Landsberg, J., Zhang, W., Clark, H., & Campbell, B. D. (2007). Plant trait responses to grazing – a global synthesis. Global Change Biology, 13(2), 313-341. doi:10.1111/j.1365-2486.2006.01288.x
- Dörner, R. P. (2023). The effects of management practices and environmental variables on the biodiversity of Nardus grasslands in the national park Hohe Tauern, Austria [Master’s thesis in environmental science, Swedish University of Agricultural Sciences]. https://stud.epsilon.slu.se/19631/
- Dumont, B., Farruggia, A., Garel, J.-P., Bachelard, P., Boitier, E., & Frain, M. (2009). How does grazing intensity influence the diversity of plants and insects in a species-rich upland grassland on basalt soils? Grass and Forage Science, 64(1), 92-105. doi:10.1111/j.1365-2494.2008.00674.x
- Farruggia, A., Dumont, B., Scohier, A., Leroy, T., Pradel, P., & Garel, J.-P. (2012). An alternative rotational stocking management designed to favour butterflies in permanent grasslands. Grass and Forage Science, 67(1), 136-149. doi:10.1111/j.1365-2494.2011.00829.x
- Gaujour, E., Amiaud, B., Mignolet, C., & Plantureux, S. (2012). Factors and processes affecting plant biodiversity in permanent grasslands. A review. Agronomy for Sustainable Development, 32(1), 133-160. doi:10.1007/s13593-011-0015-3
- Gazol, A., Tamme, R., Takkis, K., Kasari, L., Saar, L., Helm, A., & Pärtel, M. (2012). Landscape‐ and small‐scale determinants of grassland species diversity: Direct and indirect influences. Ecography, 35(10), 944-951. doi:10.1111/j.1600-0587.2012.07627.x
- Grime, J. P. (1973). Competitive exclusion in herbaceous vegetation. Nature, 242(5396), 344-347. https://www.nature.com/articles/242344a0.pdf
- Guggenberger, T., Blaschka, A., Huber, R., Schaumberger, A., Gappmaier S., Klingler A., & Unterweger, P. (2021). +2 °C: Klimaveränderung im Almgebiet. HBLFA Raumberg-Gumpenstein. https://raumberg-gumpenstein.at/jdownloads/FODOK/2021/fodok_1_25144_klimaerwaermung_alm_guggenberger.pdf
- Hejcman, M., Hejcmanova, P., Pavlu, V., & Beneš, J. (2013). Origin and history of grasslands in Central Europe – a review. Grass and Forage Science, 68(3), 345-363. doi:10.1111/gfs.12066
- Homburger, H., Lüscher, A., Scherer-Lorenzen, M., & Schneider, M. K. (2015). Patterns of livestock activity on heterogeneous subalpine pastures reveal distinct responses to spatial autocorrelation, environment and management. Movement Ecology, 3(1), 35. doi:10.1186/s40462-015-0053-6
- Howell, H. J., Mothes, C. C., Clements, S. L., Catania, S. V., Rothermel, B. B., & Searcy, C. A. (2019). Amphibian responses to livestock use of wetlands: New empirical data and a global review. Ecological Applications, 29(8), e01976. doi:10.1002/eap.1976
- Iussig, G., Lonati, M., Probo, M., Hodge, S., & Lombardi, G. (2015). Plant species selection by goats foraging on montane semi-natural grasslands and grazable forestlands in the Italian Alps. Italian Journal of Animal Science, 14(3), 484-494. doi:10.4081/ijas.2015.3907
- Körner, C. (2021). Alpine plant life: functional plant ecology of high mountain ecosystems. Springer.
- Körner, C., & Spehn, E. M. (2024). Mountain biodiversity: A global assessment. CRC Press.
- Laiolo, P., Dondero, F., Ciliento, E., & Rolando, A. (2004). Consequences of pastoral abandonment for the structure and diversity of the alpine avifauna. Journal of Applied Ecology, 41(2), 294-304. doi:10.1111/j.0021-8901.2004.00893.x
- Le Hénaff, P.-M., Galliot, J.-N., Le Gloanec, V., & Ragache, Q. (2021). Végétations agropastorales du Massif central : Catalogue phytosociologique des milieux ouverts herbacés. Conservatoire botanique national du Massif central. https://projets.cbnmc.fr/prairies/actions/catalogue-vegetations-agropastorales-massif-central/
- MacDonald, D., Crabtree, J. R., Wiesinger, G., Dax, T., Stamou, N., Fleury, P., Gutierrez Lazpita, J., & Gibon, A. (2000). Agricultural abandonment in mountain areas of Europe: Environmental consequences and policy response. Journal of Environmental Management, 59(1), 47-69. doi:10.1006/jema.1999.0335
- Marini, L., Scotton, M., Klimek, S., Isselstein, J., & Pecile, A. (2007). Effects of local factors on plant species richness and composition of Alpine meadows. Agriculture, Ecosystems & Environment, 119(3), 281-288. https://doi.org/10.1016/j.agee.2006.07.015
- Mariotte, P., Buttler, A., Johnson, D., Thébault, A., & Vandenberghe, C. (2012). Exclusion of root competition increases competitive abilities of subordinate plant species through root-shoot interactions. Journal of Vegetation Science, 23(6), 1148-1158. doi:10.1111/j.1654-1103.2012.01432.x
- Mariotte, P., Buttler, A., Kohler, F., Gilgen, A. K., & Spiegelberger, T. (2013). How do subordinate and dominant species in semi-natural mountain grasslands relate to productivity and land-use change? Basic and Applied Ecology, 14(3), 217-224. doi:10.1016/j.baae.2013.02.003
- Masé, G. (2005). The management of dry grassland in Switzerland. A Swiss federal program and its local practical application. Biotechnology, Agronomy, Society and Environment, 9(2), 133-138. https://popups.uliege.be/1780-4507/index.php?id=1551/index.php?id=17369&file=1&pid=1551&lang=fr
- Meuret, M., & Provenza, F. (2015). How French shepherds create meal sequences to stimulate intake and optimise use of forage diversity on rangeland. Animal Production Science, 55(3), 309-318. doi:10.1071/AN14415
- Napoleone, F., Probo, M., Mariotte, P., Enri, S. R., Lonati, M., Argenti, G., & Burrascano, S. (2022). Agri-environmental payments drive the conservation and forage value of semi-natural grasslands by modifying fine-scale grazing intensity. Biological Conservation, 269, 109531. doi:10.1016/j.biocon.2022.109531
- Pauler, C. M., Isselstein, J., Braunbeck, T., & Schneider, M. K. (2019). Influence of Highland and production-oriented cattle breeds on pasture vegetation: A pairwise assessment across broad environmental gradients. Agriculture, Ecosystems & Environment, 284, 106585. https://doi.org/10.1016/j.agee.2019.106585
- Pauler, C. M., Isselstein, J., Suter, M., Berard, J., Braunbeck, T., & Schneider, M. K. (2020a). Choosy grazers: Influence of plant traits on forage selection by three cattle breeds. Functional Ecology, 34(5), 980-992. doi:10.1111/1365-2435.13542
- Pauler, C. M., Isselstein, J., Berard, J., Braunbeck, T., & Schneider, M. K. (2020b). Grazing allometry: Anatomy, movement, and foraging behavior of three cattle breeds of different productivity. Frontiers in Veterinary Science, 7, 494. doi:10.3389/fvets.2020.00494
- Pauler, C. M., Zehnder, T., Staudinger, M., Lüscher, A., Kreuzer, M., Berard, J., & Schneider, M. K. (2022). Thinning the thickets: Foraging of hardy cattle, sheep and goats in green alder shrubs. Journal of Applied Ecology, 59(5), 1394-1405. doi:10.1111/1365-2664.14156
- Perotti, E., Probo, M., Pittarello, M., Lonati, M., & Lombardi, G. (2018). A 5-year rotational grazing changes the botanical composition of sub-alpine and alpine grasslands. Applied Vegetation Science, 21(4), 647-657. doi:10.1111/avsc.12389
- Pittarello, M., Probo, M., Lonati, M., & Lombardi, G. (2016). Restoration of sub-alpine shrub-encroached grasslands through pastoral practices: Effects on vegetation structure and botanical composition. Applied Vegetation Science, 19(3), 381-390. doi:10.1111/avsc.12222
- Pittarello, M., Lonati, M., Gorlier, A., Perotti, E., Probo, M., & Lombardi, G. (2018). Plant diversity and pastoral value in alpine pastures are maximized at different nutrient indicator values. Ecological Indicators, 85(Suppl. C), 518-524. doi:10.1016/j.ecolind.2017.10.064
- Pornaro, C., Schneider, M. K., Leinauer, B., & Macolino, S. (2017). Above- and belowground patterns in a subalpine grassland-shrub mosaic. Plant Biosystems - An International Journal Dealing with All Aspects of Plant Biology, 151(3), 493-503. doi:10.1080/11263504.2016.1187679
- Probo, M., Massolo, A., Lonati, M., Bailey, D. W., Gorlier, A., Maurino, L., & Lombardi, G. (2013). Use of mineral mix supplements to modify the grazing patterns by cattle for the restoration of sub-alpine and alpine shrub-encroached grasslands. The Rangeland Journal, 35(1), 85-93. doi:10.1071/RJ12108
- Probo, M., Lonati, M., Pittarello, M., Bailey, D. W., Garbarino, M., Gorlier, A., & Lombardi, G. (2014). Implementation of a rotational grazing system with large paddocks changes the distribution of grazing cattle in the south-western Italian Alps. The Rangeland Journal, 36(5), 445-458. doi:10.1071/RJ14043
- Probo, M., Pittarello, M., Lonati, M., & Lombardi, G. (2016). Targeted grazing for the restoration of sub-alpine shrub-encroached grasslands. Italian Journal of Agronomy, 11(4), 268-272. doi:10.4081/ija.2016.775
- Probo, M., Fuchs P., Schneider M. K., Hervault P., Umstätter C., Bruckmeier R. M., Pauler C. M. (2024a). Are virtual fencing applicable in mountain pastures? In C. W. Klootwijk, M. Bruinenberg, M. Cougnon, N. J. Hoekstra, R. Ripoll-Bosch, S. Schelfhout, R. L. M. Schils, T. Vanden Nest, N. van Eekeren, W. Voskamp-Harkema & A. van den Pol-van Dasselaar (Eds.), Grassland Science in Europe Vol. 29 – Why grasslands? (pp. 505-507). Proceedings of the 30th General Meeting of the European Grassland Federation. https://www.europeangrassland.org/fileadmin/documents/Infos/Printed_Matter/Proceedings/EGF2024.pdf
- Probo, M., Lonati, M., Ravetto Enri, S., Mariotte, P., & Paulerc C. (2024b). Influence de la gestion pastorale sur la conservation de la biodiversité des pâturages de montagne [Conférence]. 27e Rencontres autour des Recherches sur les Ruminants, Paris. https://journees3r.fr/recueils-2024-lintegralite-des-textes/
- Rahbek, C., Borregaard, M. K., Colwell, R. K., Dalsgaard, B., Holt, B. G., Morueta-Holme, N., Nogues-Bravo, D., Whittaker, R. J., & Fjeldså, J. (2019). Humboldt’s enigma: What causes global patterns of mountain biodiversity? Science, 365(6458), 1108-1113. doi:10.1126/science.aax0149
- Ravetto Enri, S., Probo, M., Farruggia, A., Lanore, L., Blanchetete, A., & Dumont, B. (2017). A biodiversity-friendly rotational grazing system enhancing flower-visiting insect assemblages while maintaining animal and grassland productivity. Agriculture, Ecosystems & Environment, 241, 1-10. https://doi.org/10.1016/j.agee.2017.02.030
- Ravetto Enri, S., Nucera, E., Lonati, M., Alberto, P. F., & Probo, M. (2020a). The biodiversity promotion areas: Effectiveness of agricultural direct payments on plant diversity conservation in the semi-natural grasslands of the Southern Swiss Alps. Biodiversity and Conservation, 29(14), 4155-4172. doi:10.1007/s10531-020-02069-4
- Ravetto Enri, S., Probo, M., Renna, M., Caro, E., Lussiana, C., Battaglini, L. M., Lombardi, G., & Lonati, M. (2020b). Temporal variations in leaf traits, chemical composition and in vitro true digestibility of four temperate fodder tree species. Animal Production Science, 60(5), 643-658. doi:10.1071/AN18771
- Russi, D., Margue, H., Oppermann, R., & Keenleyside, C. (2016). Result-based agri-environment measures: Market-based instruments, incentives or rewards? The case of Baden-Württemberg. Land Use Policy, 54, 69-77. doi:10.1016/j.landusepol.2016.01.012
- Schnyder, H., Locher, F., & Auerswald, K. (2010). Nutrient redistribution by grazing cattle drives patterns of topsoil N and P stocks in a low-input pasture ecosystem. Nutrient Cycling in Agroecosystems, 88(2), 183-195. doi:10.1007/s10705-009-9334-z
- Soliva, R., Bolliger, J., & Hunziker, M. (2010). Differences in preferences towards potential future landscapes in the Swiss Alps. Landscape Research, 35(6), 671-696. doi:10.1080/01426397.2010.519436
- Supek, Š., Pavlů, V., Ludvíková, V., Pavlů, L., Gaisler, J., & Hejcman, M. (2014). Effect of different grazing regimes on the coverage of Taraxacum spp. Under a long-term grazing experiment. In A. Hopkins, R. P. Collins, M. D. Fraser, V. R. King, D. C. Lloyd, J. M. Moorby & P. R. H. Robson (Eds.), Grassland Science in Europe Vol. 19 – EGF at 50:The Future of European Grasslands (pp. 324). Proceedings of the 25th General Meeting of the European Grassland Federation. https://www.europeangrassland.org/fileadmin/documents/Infos/Printed_Matter/Proceedings/EGF2014.pdf
- Svensk, M., Nota, G., Mariotte, P., Pittarello, M., Barberis, D., Lonati, M., Allan, E., Perotti, E., & Probo, M. (2022). Use of molasses-based blocks to modify grazing patterns and increase highland cattle impacts on Alnus viridis-encroached pastures. Frontiers in Ecology and Evolution, 10, 849809. doi:10.3389/fevo.2022.849809
- Svensk, M., Pittarello, M., Mariotte, P., Nota, G., Schneider, M. K., Frund, D., Dubois, S., Allan, E., & Probo, M. (2023). Nitrogen translocation by Highland cattle grazing in Alnus viridis-encroached pastures. Nutrient Cycling in Agroecosystems, 126(1), 127-141. doi:10.1007/s10705-023-10282-0
- Tasser, E., & Tappeiner, U. (2002). Impact of land use changes on mountain vegetation. Applied Vegetation Science, 5(2), 173-184. doi:10.1111/j.1654-109X.2002.tb00547.x
- Ubach, A., Guardiola, M., Oliver, X., Lockwood, M., Artola, J., & Stefanescu, C. (2023). Spatial gradients and grazing effects of a transhumant herd on plants and insect herbivores in Pyrenean subalpine grasslands. Journal of Insect Conservation, 27(5), 767-779. doi:10.1007/s10841-023-00496-6
- Wilson, J. B., Peet, R. K., Dengler, J., & Pärtel, M. (2012). Plant species richness: The world records. Journal of Vegetation Science, 23(4), 796-802. doi:10.1111/j.1654-1103.2012.01400.x
- Wood, G. M. (1987). Animals for biological brush control. Agronomy Journal, 79(2), 319-321. doi:10.2134/agronj1987.00021962007900020028x
- Zehnder, T., Lüscher, A., Ritzmann, C., Pauler, C. M., Berard, J., Kreuzer, M., & Schneider, M. K. (2020). Dominant shrub species are a strong predictor of plant species diversity along subalpine pasture-shrub transects. Alpine Botany, 130(2), 141-156. doi:10.1007/s00035-020-00241-8
Abstract
Mountain pastures are among the most biodiverse ecosystems. Moreover, they provide feed, offer essential regulating ecosystem services to society (e.g., soil carbon storage, water purification, wildfire prevention) and are the most valuable for tourism. Mountain agroecosystems are characterized by a wide small-scale variability of abiotic factors, such as altitude, aspect, slope, and bedrock. However, they are not natural, but the result of human pastoral activity carried out for millennia. Traditional management has utilized the different feeding behaviors of a variety of livestock species and breeds, as well as their specific impact on vegetation, such as selective defoliation, trampling, nutrient redistribution and seed transport. Site-adapted animal selection and herd management techniques, stocking rates, grazing periods and frequencies have been adapted to ensure a sustainable use of fodder resource and thereby have unintentionally created a habitat of outstanding biodiversity. In recent decades, traditional management has been challenged by socio-economic and environmental changes, direct payment policies influence farmer decisions, an increasing pressure of large predators complicates grazing management, and climate change alters pasture yield and its seasonal growth. In face of these challenges, farmers should redefine what site-adapted management means. Thereby, they must avoid pasture over- and underuse. In underused areas, woody species encroachment on the one hand, leads to a loss of pastureland, biodiversity and appealing landscape. Management intensification, on the other hand, causes erosion, nutrient leaching and decreases biodiversity as well. However, new technologies, enhanced knowledge transfer and result-oriented policies have the potential to promote new site-adapted management strategies to maintain mountain pasture ecosystems and their biodiversity.
Attachments
No supporting information for this articleArticle statistics
 Views: 4260
Views: 4260
Downloads
 PDF: 654
PDF: 654
 XML: 17
XML: 17