

Comment les volailles perçoivent et interprètent leur environnement : recherche scientifique et exemples d’applications
Chapeau
La science s’intéresse de longue date aux pratiques d’élevage et à la notion de sensibilité animale. En accord avec ces questionnements, il existe une demande sociétale croissante pour l’amélioration des conditions de vie des animaux de rente. Cette amélioration nécessite des connaissances de leurs capacités cognitives, qui permettent à l’animal de traiter, mémoriser, interpréter ce qui se passe dans son environnement. Ces connaissances permettent d’expliquer voire limiter certains comportements observés en élevage
Introduction
En 2016 puis en 2018, deux expertises scientifiques consécutives, l'une menée par l'autorité européenne de sécurité des aliments (European Food Safety Authority ; Le Neindre et al., 2017), l'autre pilotée par l'agence nationale de sécurité sanitaire de l'alimentation, de l'environnement et du travail (ANSES, 2018) ont abouti à une nouvelle définition du bien-être animal. Ainsi, le bien-être d'un animal correspond à « l'état mental et physique positif lié à la satisfaction de ses besoins physiologiques et comportementaux, ainsi que de ses attentes*
La santé mentale d’un animal, et donc son bien-être, dépend directement de la manière dont il est capable de percevoir, d’interpréter et d’analyser son environnement et les évènements qui s’y déroulent. Il est donc nécessaire de s’intéresser de près aux capacités mentales des animaux pour mieux appréhender les questions de bien-être relatives à chaque espèce. En effet, cette notion de santé mentale n’est pas équivalente à celle de l’humain mais est propre et particulière à chaque espèce animale. Les connaissances scientifiques actuelles permettent aujourd’hui de dire que les animaux d’élevage, comme les poules et poulets domestiques, possèdent des capacités mentales, dites capacités cognitives*, qui leur sont propres . Ces capacités cognitives regroupent un ensemble de processus mentaux* (attention*, mémorisation*, raisonnement*...) que l’animal utilise pour comprendre et s’adapter à son milieu de vie. L’objectif de cette synthèse est de faire un état des lieux non exhaustif des connaissances sur les capacités cognitives des volailles, en se focalisant sur les capacités de mémoire et d’apprentissage*. Nous nous concentrerons sur des études menées chez la poule domestique Gallus gallus domesticus (poussins, poulets de chair et poules pondeuses), qui en plus d’être les animaux de rente les plus élevés dans le monde (FAO, 2010) sont de loin les volailles les plus étudiées en cognition. Puis nous montrerons que ces connaissances peuvent être utiles pour mieux comprendre certains comportements qu’expriment les animaux dans leur milieu d’élevage.
1. Quelques notions de cognition
La cognition est définie comme l’ensemble des processus mentaux qui se rapportent aux modalités de mémorisation et de traitement des informations externes (provenant de l’environnement) et internes (physiologie et/ou ressenti) (Vauclair & Kreutzer, 2004). Une analyse récente de la littérature a montré que près de 200 articles scientifiques publiés entre 1967 et 2017 ont concerné les émotions* ou la cognition* des oiseaux domestiques comme la poule domestique (Freire & Hazel, 2017). Nous disposons donc de connaissances importantes sur leurs capacités cognitives (encadré 1). Dans cette première partie, nous reprendrons de façon synthétique les données de la littérature sur la cognition de ces oiseaux en nous focalisant sur les poules domestiques. Ces études illustrent qu’ils partagent avec bien d’autres animaux des capacités sensorielles et cognitives simples (exemple : conditionnements*, tels que : la survenue d’un son précède une récompense). De façon plus surprenante et moins connue, elles révèlent qu’ils sont aussi dotés d’un certain nombre de capacités cognitives relativement sophistiquées, comme des capacités de mémoire complexes (exemple : mémoire relationnelle, partie 1.4). Nous différencierons ici les capacités cognitives dites « simples » des capacités dites « complexes » en fonction des informations que l’animal se représente mentalement (nombre, type) et que l’animal met en relation les unes avec les autres, afin de réagir à une situation donnée. Dans un premier temps, nous ferons le point sur les capacités sensorielles des volailles, qui portent l’information environnante à la connaissance de l’animal qui peut ensuite y réagir en conséquence. Puis nous nous attarderons sur certaines capacités cognitives découvertes chez la poule domestique, en couvrant la cognition sociale, relative aux relations entre les congénères, et la cognition physique, relative à l’environnement inanimé (figure 1).
Figure 1. Exemples de capacités cognitives relatives à la cognition sociale (relative aux relations sociales) et/ou à la cognition physique (relative aux objets inanimés) mentionnés dans la synthèse.
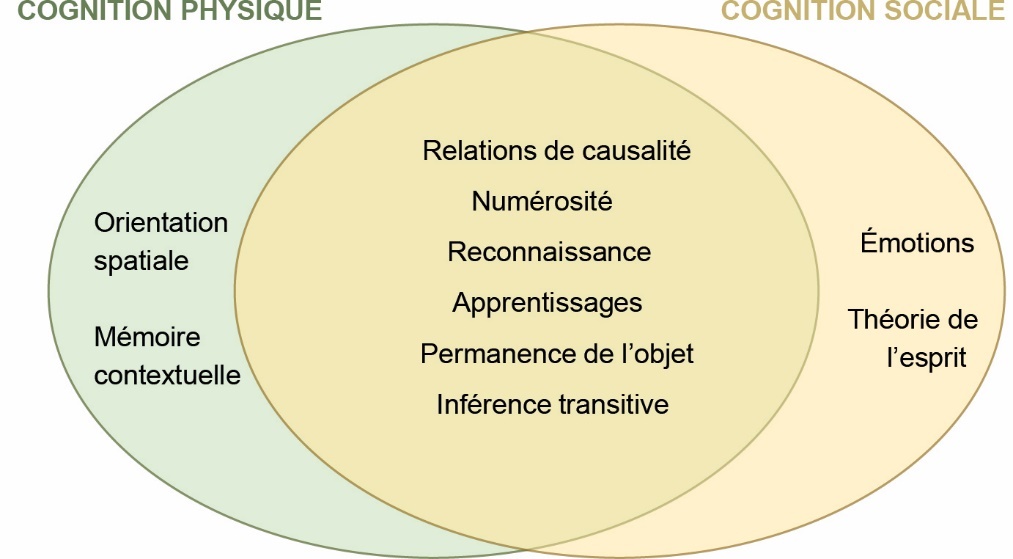
Beaucoup de capacités cognitives, comme par exemple la compréhension des relations de causalité (comprendre quelle cause a entrainé quel effet), peuvent être appliquées aux deux types de cognition. Au contraire, le développement de certaines capacités cognitives sera favorisé par un monde physique ou par un monde social complexe.
Encadré 1. Qu’est-ce que l’étude de la cognition animale ?
La cognition animale (ou éthologie cognitive) est une discipline scientifique qui consiste à étudier et comprendre le fonctionnement des processus mentaux sous-jacents aux comportements animaux. Pour étudier ces processus mentaux, les chercheurs mettent en place des tests de comportement spécifiques. Par exemple, l’animal peut être placé dans une arène de test ou face à un écran, et être exposé à une tâche afin d’observer sa capacité à la résoudre spontanément. De nombreux tests de cognition impliquent des apprentissages afin d’analyser les capacités de mémoire de l’animal, par exemple en fonction de la quantité d’information à mémoriser, ou encore sa capacité de mémorisation sur le long terme. D’autres tests mettent l’animal face à une situation nouvelle afin d’analyser les stratégies cognitives utilisées.
Tous ces tests se basent sur les capacités sensorielles et motrices de l’espèce étudiée. Connaître les capacités sensorielles et motrices de l’animal permet de réaliser des tests de cognition qui sont adaptés à sa biologie, en utilisant des signaux qu’il est capable de percevoir (par exemple : quelle gamme de fréquence de sons entend-il ?) et en plaçant l’animal dans un environnement avec lequel il sera capable d’interagir. Il est notamment important d’élaborer un dispositif de test impliquant une situation dans laquelle l’animal sera motivé à participer, soit spontanément, soit par la mise en jeu de récompenses.
Lorsqu’ils sont bien adaptés à l’espèce étudiée, ces différents tests permettent de questionner un processus cognitif bien ciblé, ce qui est souvent impossible par la seule observation des animaux dans leur milieu. D’autres approches peuvent être couplées aux tests de comportements, telles que des études de physiologie ou de neurobiologie, notamment pour mettre en évidence les régions cérébrales impliquées dans les capacités cognitives étudiées.
1.1. Les volailles, comme bien d’autres oiseaux, ont des capacités sensorielles développées
Il est maintenant admis que tous les oiseaux domestiques possèdent des capacités visuelles, auditives, olfactives et tactiles bien développées (Nicol, 2015 ; Marino, 2017 ; Garnham & Løvlie, 2018). Leur acuité visuelle est remarquable et joue un rôle primordial dans leur rapport à l'environnement (figure 2). Comme la plupart des oiseaux, ils sont capables de voir les couleurs que nous voyons (spectre de la lumière visible) et sont aussi capables de voir les fréquences lumineuses de l'ultraviolet. Ils perçoivent des fréquences lumineuses plus élevées que celles que l'œil humain est capable de voir, et vont, par exemple, percevoir l'éclairage d'un néon comme celui d'un stroboscope. En outre, chacun de leurs yeux ont un rôle plus spécifique : par exemple, ils utilisent l'œil droit pour les petits détails alors que l'œil gauche est davantage utilisé pour les informations nouvelles ou pour repérer les prédateurs. Cette capacité visuelle, qui est due à une forte latéralisation* du cerveau (i.e. séparation des fonctions entre les deux hémisphères), leur permet de porter une attention égale sur deux tâches différentes au même moment. Ils peuvent ainsi rechercher de la nourriture dans leur environnement tout en surveillant la présence éventuelle de prédateur. Après la vue, l'ouïe est le deuxième sens le plus important chez les oiseaux. Leur gamme auditive complète est similaire à la nôtre et ils perçoivent aussi les infrasons (sons plus graves) que les humains n'entendent pas. Parmi les autres sens, les poules domestiques ont aussi un sens olfactif performant. De nombreuses études ont même montré que les poussins sont capables de percevoir les odeurs in ovo et que cette expérience peut influencer leurs choix à la naissance (Bertin et al., 2010). Enfin, pour le sens du toucher, elles possèdent des capteurs nerveux essentiellement sur les pattes, le bec et la langue, ainsi qu'au niveau de petites plumes sensibles autour des yeux et du bec (Iwaniuk & Wylie, 2020) (figure 3). En plus de ces sens, les oiseaux domestiques seraient capables de s'orienter dans l'environnement en détectant les champs magnétiques, grâce à des composants présents dans leur rétine, les cryptochromes, à l'instar des pigeons (Freire et al., 2005 ; Wiltschko et al., 2021).
Compte tenu de ces connaissances, il apparaît que les oiseaux domestiques évoluent dans un univers sensoriel riche et diversifié. Le fait qu’ils possèdent tous ces sens fonctionnels laisse également penser qu’ils ont les capacités pour pouvoir traiter, mémoriser voire interpréter tout ou partie des informations sensorielles perçues dans leur environnement social (groupe de congénères) et physique (milieu de vie).
Figure 2. Les capacités sensorielles de la poule sont la première entrée qui lui permet d’appréhender le monde qui l’entoure.

Les informations provenant de l’environnement sont traitées, puis interprétées par l’animal en fonction de ses capacités cognitives. Chez la poule, l’exploration visuelle est prépondérante lors de la rencontre avec un nouvel objet. (Photo : Rachel Degrande)
Figure 3. Photographie montrant les petites plumes sensibles entourant l’œil et le bec de la poule.

(Photo : libre de droits, 1ZOOM.ME).
1.2. Les poules domestiques sont des animaux sociaux qui possèdent des capacités cognitives sociales
La cognition sociale concerne les capacités cognitives relatives aux interactions entre un individu et les différents membres du groupe. Elle comprend la perception et le traitement des informations sociales par rapport au comportement des congénères. Les capacités cognitives sociales des oiseaux domestiques ont essentiellement été étudiées chez la poule domestique. À l'état sauvage comme à l'état domestique, les poules sont des animaux sociaux qui vivent en groupe, au sein d'une structure sociale complexe et hiérarchisée. Afin de pouvoir évoluer au sein d'une telle structure, elles ont développé des capacités spécialisées (Bradshaw & Dawkins, 1993 ; Nicol, 2015). Dès leur plus jeune âge, les poussins sont capables de se servir des informations sociales pour s'adapter à leur environnement via une capacité d'apprentissage social. Ils sont, par exemple, capables d'apprendre quel type de nourriture a un effet néfaste en observant la réaction d'un congénère qui y a goûté (Salva et al., 2009), ce que font aussi les poussins de caille (Akins & Zentall, 1998). La mère a aussi une part importante dans les apprentissages des poussins : lorsque les poussins picorent une nourriture que la poule a associée avec une expérience négative (aliment au goût amer manipulé expérimentalement), la mère gratte le sol et émet des vocalisations particulières, ce qui éloigne les poussins de la nourriture considérée néfaste et les incite à rechercher de la nourriture au sol proche d'elle.
Un autre exemple est leur capacité à reconnaître et à distinguer l'identité des congénères de leur groupe et leur statut hiérarchique* relatif (poule : Hogue et al., 1996 ; oie cendrée : Weiß et al., 2010), mais aussi de catégoriser* si un individu appartient à un groupe social étranger (Bradshaw & Dawkins, 1993). Ces capacités de discrimination ont été démontrées en conditions naturelles mais aussi en conditions contrôlées, en utilisant des stimuli artificiels (par exemple : photographies de congénères) et selon différents canaux sensoriels (ouïe, vue : DeMello et al., 1993). Cela leur permet notamment de développer des affinités à l'égard de certains individus du groupe.
L'étude des vocalisations des poules domestiques a également permis de mettre en évidence leur capacité à communiquer et interagir. Les poules sont capables d'adapter leurs vocalisations en fonction de la situation, des individus présents ou encore de l'âge de leurs petits (par exemple : Karakashian et al., 1988). À titre d'exemple, elles émettent des vocalisations différentes selon que le prédateur rencontré soit un prédateur terrestre ou aérien (Marler et al., 1987). Il a aussi été observé que des coqs sont capables, en fonction de la présence ou non du coq dominant, d'adapter leur comportement et leurs vocalisations pour ne pas se faire repérer lorsqu'ils cherchent à attirer des femelles. Cette prise en compte de l'autre et de son statut (ici, le coq dominant) est une capacité associée chez certaines espèces à la théorie de l'esprit* (représentation mentale de ce que l'autre peut percevoir ou non). Les vocalisations émises au sein du groupe font notamment partie des indices comportementaux perçus par les congénères et peuvent jouer un rôle dans la contagion émotionnelle* sociale. Par exemple, un individu qui perçoit un stress chez des congénères va émettre en retour ces mêmes comportements ou indices physiologiques de stress (Edgar et al., 2013). Plus encore, une poule mère qui a vécu un évènement négatif dans une certaine situation va émettre des comportements de stress lorsque ses poussins sont exposés à cette même situation (Edgar et al., 2011). Ces résultats suggèrent que non seulement la poule est sujette à la contagion émotionnelle face au comportement de congénères, mais qu'elle serait aussi capable d'utiliser sa mémoire des évènements pour montrer un comportement s'apparentant à une forme d'empathie*.
1.3. Les poules domestiques possèdent des capacités d’apprentissage et de mémoire
L'une des capacités de mémoire les plus connues chez les oiseaux domestiques est l'empreinte*. L'empreinte est une forme de mémoire spécifique qui permet aux jeunes de mémoriser « le premier objet mobile » qu'ils voient dans leur environnement dans les heures qui suivent l'éclosion (Nakamori et al., 2013). L'« objet mobile » en question peut être leur mère ou un objet en particulier que l'on aura mis en présence des poussins lors de la période sensible*. Le phénomène d'empreinte est tel que si l'objet se déplace, le poussin le suivra automatiquement et exprimera une préférence pour cet objet. Cette mémoire n'implique pas de processus mentaux sophistiqués et s'établit automatiquement. Toutefois, c'est une mémoire qui a la particularité de s'établir très rapidement et d'être très robuste à l'oubli (McCabe, 2019).
Les oiseaux domestiques comme la poule sont capables d'apprentissages associatifs* : ils sont capables d'associer rapidement une localisation, un objet ou même une action à la survenue d'un évènement positif ou négatif (exemple chez l'oie cendrée : Fritz et al., 2000 ; chez la poule domestique : Martin & Bellingham, 1979). Ces apprentissages peuvent être flexibles et reproduits en conditions contrôlées* (Degrande et al., 2022). Par exemple, une poule apprendra très rapidement que piquer sur un bouton rouge lui permet d'obtenir une récompense, alors que le bouton vert présenté en même temps ne lui donne rien. Si la demande change, c'est-à-dire si la récompense est cette fois délivrée lorsqu'elle pique sur le vert, alors la réponse comportementale de la poule changera en conséquence, et ce très rapidement (figure 4a). Cette flexibilité comportementale*, qui permet à l'animal de s'adapter rapidement à des situations changeantes, peut être influencée par des facteurs individuels comme la personnalité* des individus (i.e., traits de caractère propres à l'individu et persistants, comme l'anxiété ou la témérité). Les apprentissages associatifs peuvent aussi permettre de questionner d'autres capacités cognitives chez les animaux, comme leur capacité à comprendre les intervalles de temps (i.e., durée entre deux évènements) ou encore à anticiper la survenue d'une récompense à intervalle fixe, ce qui a été montré chez la poule (Zimmerman et al., 2011).
Les poules domestiques possèdent bien d'autres formes de mémoire. Des chercheurs ont ainsi montré que ces oiseaux possèdent une mémoire spatiale très performante (figure 4b). Il s'agit de la capacité à mémoriser les informations spatiales (les différents objets et leur emplacement) qui composent leur environnement. Il a par exemple été montré que le poussin âgé de quelques jours (Vallortigara et al., 1990), comme des individus plus âgés, sont capables de se repérer dans l'espace grâce à deux mémoires différentes (aussi montré chez la caille : Lormant et al., 2020). D'un côté, ils peuvent apprendre à se repérer dans leur environnement en utilisant une mémoire relativement simple dite « indicée », basée sur quelques repères saillants de leur environnement. Autrement dit, l'animal se repère dans son environnement grâce à la reconnaissance d'éléments particuliers de l'environnement (exemple : la mangeoire est à droite de l'arbre). D'un autre côté, ils sont aussi capables d'utiliser une mémoire plus complexe et plus flexible, basée sur une représentation mentale riche, qui consiste en une carte mentale des nombreux éléments qui composent leur environnement. Dans ce cas, l'animal se repère dans son environnement grâce à la relation spatiale relative entre les éléments de l'environnement (exemple : la mangeoire se trouve entre l'arbre et le poulailler). Ces études montrent ainsi que les oiseaux domestiques ne possèdent pas seulement des capacités cognitives simples, ils semblent aussi dotés de capacités cognitives plus sophistiquées.
Si beaucoup d’études se sont servies des capacités d’apprentissages associatifs et des spécialisations adaptatives* des poules pour en apprendre plus sur leur monde mental, les capacités dont nous avons discuté peuvent être qualifiées de « simples » car ne mettent pas nécessairement en œuvre une forme de raisonnement (manipulation d’images mentales, ou d’informations apprises, en lien avec un objectif). Plus récemment, les capacités d’apprentissage de la poule domestique ont poussé les chercheurs à s’intéresser de plus près à des capacités plus élaborées, dont certaines que l’on retrouve chez les oiseaux considérés intelligents comme les corvidés, et qui impliquent des capacités cognitives que l’on pensait propres à l’humain.
1.4. Les poules domestiques sont capables de cognition élaborée
En utilisant le phénomène d'empreinte mentionné précédemment, des recherches ont montré que les oiseaux domestiques possédaient des capacités cognitives plus étonnantes. Ainsi, le poussin âgé de quelques jours comprend la notion de permanence de l'objet. Cette capacité cognitive permet d'intégrer le fait qu'un objet existe même s'il n'est plus dans le champ de vision de l'individu, ou encore de suivre un objet en mouvement et déduire sa trajectoire probable. Plus encore, cette capacité permet de reconnaître un objet ou une forme géométrique, même si celui-ci est partiellement occulté ou si l'on ne peut en voir que les contours subjectifs (i.e., perception de contours alors qu'ils ne sont pas explicitement dessinés, illusion d'optique) (Zanforlin, 1981 ; Regolin & Vallortigara, 1995 ; Lea et al., 1996). Le jeune poussin possède également une compréhension rudimentaire de la cohérence physique des objets : les poussins montrent une préférence spontanée pour des images représentant des objets possibles, plutôt que des images dont l'information structurelle est inconsistante (Regolin et al., 2011).
Parmi les capacités les plus étonnantes découvertes chez la poule, une équipe de chercheurs a montré que des poussins, en plus d'être capables de discriminer des quantités, pouvaient réaliser des opérations de calcul mental (Rugani et al., 2009). Dans cette expérience, c'est le déplacement d'objets sur lesquels le poussin a dirigé son empreinte qui est utilisé : deux panneaux opaques sont face au poussin, et l'expérimentateur place, à la vue du poussin, une certaine quantité de ces objets derrière chacun des panneaux (à gauche ou à droite ; figure 4c). Le poussin se dirige automatiquement vers le panneau qui cache le plus d'objets. Puis lorsque l'expérimentateur déplace un nombre déterminé d'objets d'un écran à l'autre, le poussin réussit à déduire derrière quel écran se cachent le plus d'objets et ce indépendamment de la direction du flux, ce qui n'est possible que par additions et soustractions. La même équipe de chercheurs a remarqué que le poussin classe mentalement les plus petites quantités à gauche et les plus grandes quantités à droite ; ce qui veut dire qu'il possède une ligne mentale numérique (Rugani et al., 2015) comme celle présente chez plusieurs mammifères dont l'humain.
Figure 4. Différents dispositifs de test en cognition animale chez les volailles.

(a) Des dispositifs sont développés afin de tester les capacités cognitives chez la poule domestique de manière automatisée. Sur l'illustration, la poule pique sur une image qui s'affiche à l'écran pour obtenir une récompense (Degrande et al., 2022). (Photo : © Fabien Cornilleau).
(b) Cette arène de test de mémoire spatiale contient des contenants identiques mais un seul cache de la nourriture. La caille, comme la poule, est capable d'apprendre à retrouver l'emplacement du seul pot qui contient la nourriture. Pour cela les animaux mémorisent les indices spatiaux disposés sur les murs de l'arène (mémoire spatiale, Lormant et al., 2020). (Photo : © Ludovic Calandreau).
(c) Les chercheurs dirigent l'empreinte (attachement et reconnaissance précoce biologiquement dirigés vers la mère) du poussin envers un objet par exposition lors de la période sensible. Grâce à cette mémoire forte, ils montrent que le poussin réalise des opérations mentales simples et se dirige vers le panneau opaque qui contient le plus d'objets (Rugani et al., 2009). (Photo : © Université de Padoue, Université de Trente).
(d) Les poulets peuvent être équipés de ponchos souples (code d'identification placé autour du cou) ce qui permet de les identifier individuellement lorsqu'ils sont sur le parcours extérieur. Grâce aux observations, la propension de chaque individu à explorer le parcours (plus ou moins loin et plus ou moins fréquemment) peut ainsi être caractérisée. Ce trait de comportement pourra ensuite être mis en relation avec les capacités cognitives individuelles (Ferreira et al., 2020b). Photo : © Vitor Hugo Bessa Ferreira).
Des études ont également montré que le poussin est capable d'inférence transitive (Daisley et al., 2010). Cette capacité cognitive est associée à une forme de raisonnement puisqu'elle permet de comparer des informations mémorisées (mémoire relationnelle). Ainsi, si nous apprenons que A est supérieur à B et que B est supérieur à C, nous pouvons en déduire de manière indirecte que A est supérieur à C. La poule peut utiliser cette capacité pour se positionner socialement au sein d'un groupe, uniquement en observant les interactions sociales* entre les membres du groupe (Hogue et al., 1996). La capacité d'inférence transitive a aussi été démontrée chez l'oie en conditions expérimentales* (Weiß et al., 2010).
L'inférence transitive fait partie des capacités cognitives dites complexes car elle met en jeu des représentations mentales, que l'animal utilise ensuite pour adapter son comportement à une situation donnée. Dans le cas de l'inférence transitive, les représentations mentales que l'animal a mémorisées et qui sont mises en jeu sont les identités des congénères et les relations de hiérarchie qui les lient. D'autres capacités cognitives mettant en jeu des représentations mentales ont été démontrées chez la poule domestique, comme la compréhension du concept* de correspondance à l'échantillon. Dans ce type d'expérience, la poule est mise face à un stimulus dit l'échantillon (exemple : un rond de couleur rouge) et doit ensuite retrouver, parmi plusieurs (exemple : rond de couleur bleu et rond de couleur rouge), le stimulus qui correspond à l'échantillon présenté juste avant (Nakagawa et al., 2004). Un autre exemple est la mémoire contextuelle : en plus de posséder la mémoire de la localisation d'une récompense (Forkman, 2000 ; Cozzutti & Vallortigara, 2001) une étude récente montre que le poussin est capable d'associer l'emplacement de la récompense (cachée à droite ou à gauche) selon le contexte (Loconsole et al., 2021). Ces capacités sont reliées à la mémoire déclarative*, qui englobe la réutilisation explicite d'informations qui ont été mémorisées à court terme ou à long terme. Ce système de mémoire peut notamment être impliqué dans la résolution de problèmes ou la prise de décision.
Plus surprenant encore, une étude tend à démontrer que les poules seraient capables de métacognition, plus précisément de métamémoire*. Cette capacité permet à un individu « d'évaluer mentalement le niveau de ses connaissances » (par exemple : « est-ce que je connais la réponse à la question ? »), ce qui a longtemps été considéré comme propre à l'humain. Des chercheurs de l'université de Kyoto au Japon ont étudié cette capacité chez des oiseaux comme le pigeon mais aussi chez la poule domestique naine (Nakamura et al., 2011). Dans cette expérience, les oiseaux ont d'abord été entrainés, en piquant avec leur bec, à retrouver un rond coloré lorsque celui-ci apparait sur un écran parmi plusieurs autres ronds de couleurs différentes (tâche de correspondance à l'échantillon), en échange d'une récompense. Une fois cette tâche apprise, les chercheurs ont inséré une nouvelle étape. Désormais, deux symboles (un carré orange et un carré violet) apparaissent sur l'écran aussitôt que les oiseaux ont réalisé la tâche des ronds de couleur. Si l'oiseau choisit de piquer sur le symbole orange, il obtient la récompense attendue uniquement s'il a correctement répondu lors de la première tâche avec les ronds de couleur. Piquer sur ce symbole est donc une « prise de risque » : dans le cas où l'animal choisit ce symbole orange alors qu'il a donné la mauvaise réponse lors de la première tâche, il n'obtiendra aucune récompense. En revanche, si l'oiseau choisit le symbole violet il obtiendra une récompense intermédiaire, plus petite, qu'il ait ou non réussi la tâche avec les ronds de couleur. Piquer sur le symbole violet est donc associé à la sécurité, la garantie d'obtenir une petite récompense, que la réponse soit correcte ou incorrecte lors de la première tâche. De façon remarquable, les résultats montrent que les oiseaux ne répondent pas au hasard lors de cette nouvelle étape. En effet, certains d'entre eux sélectionnent davantage le symbole violet, associé à la sécurité, lorsqu'ils ont mal répondu lors de l'épreuve des ronds de couleur. En d'autres termes, ces oiseaux semblent capables d'estimer leur niveau de connaissances. Cette expérience révèle donc que les poules domestiques semblent capables de métacognition. À ce jour, cette expérience n'a pas encore été tentée par d'autres équipes de recherche chez d'autres oiseaux domestiques. Il est donc important de ne pas surinterpréter ces résultats. Néanmoins, cette étude et ses résultats invitent à se questionner sur l'étendue des capacités cognitives des oiseaux domestiques.
1.5. Quelles fonctions ont ces capacités cognitives pour l’animal ?
Cette revue synthétique des capacités sensorielles et cognitives chez les oiseaux domestiques illustre que ces oiseaux ont certainement un répertoire de capacités cognitives bien plus étendu que ce que nous pouvions imaginer. Les études qui ont permis de le démontrer ont été, pour la plupart, réalisées dans des conditions contrôlées en laboratoire, ce qui pourrait laisser penser que dans la nature ou dans les élevages ces oiseaux ne disposent pas de telles capacités cognitives ou ne les utilisent pas. Mais il faut nuancer cette idée : les expériences menées ne créent pas les capacités cognitives chez les animaux testés, elles contribuent en revanche à révéler leur existence en posant les questions de façon appropriée. Ces études démontrent clairement que les oiseaux domestiques n’ont pas uniquement des réponses-réflexe. Ils disposent d’un univers mental qui leur est propre, sont capables d’apprentissages et savent réutiliser des informations qu’ils ont en mémoire. Pour autant, il ne faut pas non plus surinterpréter les résultats obtenus dans les expériences de cognition. Il est évident que les oiseaux domestiques ne sont pas de grands mathématiciens ou encore de grands philosophes. En revanche, les poussins sont probablement capables de discriminer les quantités et réaliser des calculs sur de petits nombres car il est important pour les jeunes d’être capables de rejoindre le groupe d’individus composé du plus grand nombre. En effet, un poussin de quelques jours ne peut pas encore réguler sa température corporelle. Rejoindre le groupe composé du plus grand nombre lui garantira davantage de profiter de la chaleur des autres. Par ailleurs, un poussin aura moins de chance d’être attrapé par un prédateur s’il est situé au milieu d’un grand groupe de congénères. Enfin, les capacités de métacognition évoquées plus tôt leur permettraient d’émettre des réponses en situation de risque ou de gérer des situations d’incertitudes qui peuvent survenir. Les capacités cognitives sont essentielles aux animaux pour traiter, mémoriser les informations et événements auxquels ils sont confrontés. Elles constituent des éléments indispensables à leur adaptation, notamment en élevage.
2. L’intérêt des études de cognition pour l’élevage
Dans la première partie de cette synthèse nous avons montré que les oiseaux domestiques possédaient un certain nombre de capacités cognitives qui leur sont utiles pour traiter et mémoriser les informations et événements auxquels ils sont confrontés. Dans cette seconde partie, nous illustrerons en quoi les études de cognition peuvent permettre de mieux comprendre certains comportements de ces oiseaux en élevage.
En effet, l’évolution de nos connaissances sur leurs capacités cognitives permet également de proposer des solutions pour enrichir ou améliorer les conditions de vie des animaux en élevage. Pour ne prendre que quelques exemples, une première application serait le développement d’outils d’évaluation de la valence* de certains stimuli pour l’animal (et ainsi de son confort dans diverses conditions), en nous basant sur nos connaissances de ses capacités sensorielles couplées à nos connaissances sur le ressenti individuel (par exemple : évaluation du comportement de l’animal en présence de certains objets). En parallèle, l’étude des capacités d’apprentissage social chez une espèce peut permettre de mieux comprendre et empêcher la diffusion des comportements anormaux au sein du groupe, comme le picage sévère*. Enfin, dans l’environnement d’élevage, des enrichissement cognitifs associés à la survenue de nourriture pourront être proposés sur la base des capacités de mémorisation et d’anticipation des volailles.
Pour illustrer les liens entre études de cognition et comportements en élevage, nous nous focaliserons sur deux problématiques majeures : les différences interindividuelles dans l'utilisation des parcours extérieurs, et les comportements de picage sévère. Bien que plusieurs facteurs externes aient pu être mis en évidence pour expliquer en partie ces comportements (Dawkins et al., 2003 ; Bestman et al., 2019), le développement des approches cognitives permet de mieux comprendre les causes individuelles de l'apparition de ces comportements, et ainsi d'adapter les stratégies de gestion des animaux afin d'en diminuer les effets négatifs à long terme (Nicol, 2004 ; Freire, 2020 ; Ferreira et al., 2021).
2.1. Relation entre cognition et exploration du parcours
Les poulets de chair et poules pondeuses élevés en plein-air ont accès à une zone située à l'extérieur du bâtiment d'élevage, appelée le parcours (figure 5). Le parcours est considéré comme une zone d'enrichissement environnemental qui permet aux animaux d'exprimer une plus grande diversité de leur répertoire comportemental, par exemple par les bains de poussière, le grattage au sol, ou encore l'exploration de leur environnement. Plusieurs études ont montré que l'utilisation du parcours extérieur peut dépendre d'un certain nombre de facteurs externes comme la saison, le moment de la journée, la présence d'un couvert végétal ou encore des facteurs internes, comme la génétique, le sexe ou l'âge des animaux (Pettersson et al., 2016 ; Bestman et al., 2017 ; Campbell et al., 2021). Toutefois, ces facteurs n'expliquent pas pourquoi, au sein d'un même élevage, il demeure toujours une variabilité importante entre les individus dans l'utilisation du parcours extérieur. En effet, pour un même lot, certains individus exploreront très souvent les zones les plus éloignées du parcours alors que d'autres resteront à proximité de leur bâtiment d'élevage (Gebhardt-Henrich et al., 2014 ; Taylor et al., 2017a ; Taylor et al., 2017b ; Larsen et al., 2018 ; Ferreira et al., 2022). Cette différence de comportement suggère que l'utilisation du parcours peut dépendre de l'individu lui-même, notamment de la façon dont il traite et mémorise les informations de son environnement, en d'autres termes de ses capacités cognitives.
Figure 5. La tendance des individus à explorer le parcours extérieur dépend de différents facteurs.

La fréquence des visites à l’extérieur ou encore l’éloignement par rapport au bâtiment dépendent par exemple de la distribution du couvert végétal, de la saison, ou encore de facteurs liés à l’individu comme l’âge ou la personnalité. Sur cette photo, certains poulets choisis au hasard sont équipés de ponchos souples (code d’identification placé autour du cou) afin de permettre leur identification rapide lors des analyses d’exploration du parcours. (Photo : © Vitor Hugo Bessa Ferreira)
Pour tester si les animaux plus ou moins explorateurs montrent des différences en termes de capacités cognitives, des poulets ont été soumis à un test qui est fréquemment utilisé chez d'autres espèces afin d'évaluer leur capacité à s'adapter rapidement. Ce test se déroule en deux temps. Dans un premier temps, les animaux sont entrainés à contourner un tube opaque pour récupérer de la nourriture placée à l'intérieur. Dans un second temps, la même tâche est présentée, mais cette fois avec un tube transparent. Les animaux doivent alors refreiner leur envie première de picorer la paroi transparente du tube, qui est une stratégie inefficace, pour récupérer la nourriture. Ils doivent faire preuve de flexibilité et utiliser ce qu'ils ont appris lors de la première étape : contourner le tube pour récupérer la nourriture. Les résultats de cette expérience montrent que les individus les moins explorateurs du parcours obtiennent de meilleurs scores (i.e., comprennent plus rapidement et picorent peu ou pas la paroi transparente) que les individus les plus explorateurs du parcours (Ferreira et al., 2020b). En parallèle, d'autres études font le lien entre la flexibilité comportementale (i.e., la capacité à s'adapter aux situations changeantes), le comportement de vigilance, et la tendance des poules à passer plus de temps en intérieur (Sih & Del Giudice, 2012 ; Campbell et al., 2019). Le comportement de vigilance fait référence à l'attitude alerte de l'animal et peut être dû soit à un trait de personnalité (trait persistant d'anxiété plus ou moins prononcée), soit à un état émotif anxieux ponctuel dû à un évènement externe. Il est donc possible, pour les poulets les moins explorateurs, que ce trait de vigilance influence négativement l'utilisation du parcours chez certains individus. L'expérience avec les tubes transparents aura permis de mettre en évidence le lien entre certaines capacités cognitives de l'individu (ici, la flexibilité) et son comportement sur le parcours (figure 4d).
Le lien entre un poulet moins explorateur et un meilleur score au test cognitif est conforté par d'autres études, qui montrent que les poulets les moins explorateurs ont aussi de meilleures capacités de mémoire spatiale que les poulets les plus explorateurs (Ferreira et al., 2019). Des poulets ont été entraînés à retrouver de la nourriture dans une arène. La nourriture était placée uniquement dans certains bols parmi plusieurs (toujours les mêmes), différenciables par leur couleur et par leur localisation spatiale. Lors du test, les poulets ne peuvent se référer qu'à la localisation spatiale relative des bols (i.e. les bols contenant la nourriture ne sont plus identifiables par leur couleur), et doivent donc se servir des indices spatiaux de l'arène de test pour retrouver les bols contenant la nourriture (même principe que figure 4b). Les résultats montrent encore une fois que les poulets les moins explorateurs ont de meilleures performances (mémoire spatiale, relationnelle) : les individus les moins explorateurs mettent moins de temps à retrouver les bols contenant de la nourriture par rapport aux individus les plus explorateurs. Ce résultat semble contre-intuitif, dans le sens où il était possible d'imaginer que les animaux qui explorent le plus leur environnement devraient avoir une meilleure mémoire spatiale, or les résultats indiquent le contraire. Cette même conclusion est aussi observée chez d'autres espèces, oiseaux et mammifères (par exemple : Coppens et al., 2010 ; Zidar et al., 2018 ; mais conclusion contradictoire par : Dougherty & Guillette, 2018). Par ailleurs, les poulets les plus explorateurs comme les moins explorateurs sont capables d'utiliser un indice plus simple, la couleur des bols, pour retrouver la nourriture si leur localisation spatiale est modifiée (mémoire indicée, non relationnelle) (Ferreira et al., 2019). Ce dernier résultat montre l'importance de réaliser des tâches cognitives plus complexes pour différencier les poulets les plus explorateurs des moins explorateurs.
Ces études suggèrent que les poulets les moins explorateurs traiteraient plus finement les informations de leur environnement et les mémoriseraient mieux. Un tel profil cognitif pourrait guider leur décision et inhiber leur motivation à explorer un parcours extérieur. Des études similaires menées chez la poule pondeuse tendent à montrer des résultats inverses (Campbell et al., 2020). Les poules les plus exploratrices du parcours posséderaient de meilleures performances de mémoire spatiale. De plus, il apparaît que la sociabilité* et la capacité d'apprentissage social des individus peut jouer un rôle dans les comportements d'exploration des parcours (Ferreira et al., 2020a). Ces résultats soulignent que nous ne pouvons pas généraliser les résultats obtenus dans ces recherches qui sont très récentes. Toutefois, les études illustrent qu'il existe un lien entre cognition et exploration du parcours. Elles indiquent que les études de cognition pourraient, à terme, apporter de nouvelles connaissances sur la biologie de ces oiseaux, utiles pour mieux comprendre leur comportement (encadré 2). Ces connaissances pourraient permettre d'adapter au mieux leur milieu de vie pour que celui-ci couvre les besoins de tous les individus malgré leurs différences.
Encadré 2. Comment l’étude de la cognition des animaux nous permet de mieux les comprendre et d’adapter leurs conditions de vie en élevage.
Les recherches en cognition animale permettent de caractériser l’étendue des capacités cognitives des animaux. Ces études permettront d’évaluer finement comment l’animal comprend son environnement, s’il est capable de mémoriser les évènements et d’adapter son comportement en fonction des situations. Comme les capacités cognitives sont à la base de très nombreux comportements, une caractérisation complète de ces dernières permettra de mieux comprendre les processus mentaux qui peuvent être impliqués dans des comportements observés en élevage, comme l’utilisation du parcours des poulets plein-air. Ces connaissances pourraient permettre de mieux expliquer les comportements observés, leurs altérations éventuelles, ou encore permettre de mieux comprendre l’hétérogénéité dans les comportements des individus, liée à leur personnalité. Elles pourront aussi constituer une base scientifique pour, par exemple, orienter les pratiques d’enrichissement en élevage sur une mise en pratique mieux ciblée et donc plus fonctionnelle, comme les modalités d’utilisation des abris pour favoriser l’exploration du parcours extérieur.
2.2. Relation entre cognition et picage
Donner des coups de bec est un comportement naturel chez les poules. Elles picorent le sol lorsqu'elles cherchent de la nourriture ou prennent des bains de poussière, et elles se donnent également des coups de becs entre elles pour communiquer. Par exemple, des coups de becs légers peuvent avoir une valence positive dans le cadre du toilettage social. Mais le picage peut aussi être agressif lorsque les poules montrent leur dominance et établissent une hiérarchie. Cependant, lorsque le picage est fréquemment dirigé vers la peau et les plumes d'autres individus, il peut devenir un problème majeur de bien-être. Ce comportement est appelé picage sévère. Le picage sévère peut engendrer des blessures physiques lorsqu'il est effectué directement sur la peau ou au niveau du cloaque, ou encore par arrachage de plumes et exposition directe de la peau aux futurs picages. Les animaux qui subissent ce picage augmentent leur consommation alimentaire de manière à augmenter leur production de chaleur et maintenir leur température corporelle. Ceci altère directement la qualité de vie des animaux, en plus d'avoir une conséquence négative sur la production d'œufs (Cronin et al., 2020).
La complexité du picage réside dans son caractère multifactoriel (Coton et al., 2019). Jung et Knierim (2018), dans une revue récente d'études expérimentales et épidémiologiques, ont trouvé au moins 62 facteurs qui peuvent augmenter significativement le risque de développer du picage. Parmi ces facteurs se trouvent de faibles possibilités de fourragement* (le picage normalement dirigé vers l'environnement par exploration alimentaire, est alors redirigé vers les congénères) ou encore une forte densité d'animaux. Un niveau de stress élevé au sein de l'environnement de vie peut aussi être impliqué, lié notamment à des conditions environnementales frustrantes (niveau d'enrichissement par exemple), le tempérament des animaux congénères, ou encore par un stress aigu élevé causé par un évènement ponctuel (par exemple : Newberry et al., 2007 ; Van Staaveren & Harlander, 2020 ; mais conclusion contradictoire par : Pichová et al., 2021). Toutefois, des études ont démontré qu'une petite proportion d'individus semble responsable de la propagation du picage dans un groupe donné (Rodenburg et al., 2004), portant l'attention de la recherche vers la caractérisation de ces individus. Ces résultats suggèrent que le picage pourrait, en partie au moins, dépendre de l'individu et de la façon dont celui-ci perçoit son environnement et les événements qui s'y déroulent. Des études vont dans ce sens et démontrent un effet de l'apprentissage social des individus sur le développement du picage au sein du groupe. Dans une expérience, Cloutier et al. (2002) ont tenté de reproduire les conditions d'apprentissage social du picage et de les comparer à une condition sans apprentissage social (poules naïves). Les chercheurs ont exposé des jeunes poules à un individu démonstrateur*, qui a appris à piquer un modèle de poule inanimé, mimant le comportement de picage. Leurs résultats montrent que des poules, qui ont pu observer d'autres individus piquer le modèle, reproduisaient davantage ce comportement par rapport aux poules naïves. De façon similaire, Zeltner et al. (2000) ont observé que le picage se développe et augmente significativement plus dans des groupes de poussins qui contiennent quelques poussins présentant des fréquences de picage élevées.
Si le picage s'acquiert et diffuse dans le groupe en partie via un apprentissage social, d'autres études suggèrent que les poules pourraient aussi apprendre à réprimer ce comportement. Une série d'expériences confirme cette possibilité : les poules qui ont été exposées et conditionnées à des plumes trempées dans une solution de quinine (avec un goût amer, désagréable) ont montré une fréquence de picage moins élevée lorsqu'elles étaient en groupe (Harlander-Matauschek et al., 2008). De la même façon, une application de quinine sur les plumes des animaux, dès la première semaine après la naissance jusqu'à leur première ponte, réduit la fréquence des picages sévères. Cependant, une fois que le traitement a été arrêté, ces mêmes oiseaux ont montré une fréquence de picage sévère plus élevée à la période de ponte suivante, par rapport à la période de ponte précédente (Harlander-Matauschek et al., 2010). Enfin, des études de comportement, de physiologie et de neurobiologie menées sur des poules sélectionnées sur leur propension à piquer montrent que le picage sévère pourrait s'apparenter à des désordres cognitifs ou cérébraux proches de ce qui est observé dans les pathologies liées à l'hyperactivité (Kjaer, 2009). En particulier, le picage sévère mettrait en jeu des modifications du fonctionnement d'un système de neurotransmission* dans le cerveau des animaux, le système dopaminergique, qui est connu pour être impliqué dans des processus cognitifs tels que l'attention, la mémoire. Bien que certaines études tendent à montrer une absence de différence entre les poules plus ou moins sujettes au picage sévère lors de certaines tâches cognitives (self-contrôle moteur* : Heinsius et al., 2020 mais conclusion contradictoire par : Pichová et al., 2021 ; mémoire : Nordquist et al., 2011), l'effet de multiples autres capacités cognitives telles que les capacités de discrimination ou de raisonnement restent à explorer, en plus des recherches concernant l'apprentissage social. Une approche au niveau de l'individu et de sa cognition participera à élaborer des stratégies de management alternatives aux stratégies actuelles, comme l'épointage du bec.
Conclusion
Les études en cognition animale permettent d’en apprendre plus sur la manière dont les animaux comprennent leur environnement. Dans cette revue nous avons mis en lumière que les poules possèdent des capacités sensorielles variées, qu’elles sont capables d’apprentissage et de mémoire, et qu’elles possèdent une cognition plus élaborée que ce qui est souvent imaginé. À travers l’exemple de l’utilisation du parcours et des comportements de picage, nous avons ensuite montré de quelle manière la recherche en cognition peut permettre d’appréhender le vécu mental de l’animal, et ainsi participer à comprendre les problématiques rencontrées en élevage et proposer de nouvelles solutions. Nous avons aussi pu appréhender que la cognition, à elle seule, ne suffit pas à expliquer le comportement et les capacités d’adaptation de l’animal, qui dépendent aussi de facteurs individuels (personnalité, âge, composantes émotionnelles) et environnementaux (stabilité de l’environnement physique et social).
Le domaine de la cognition est un domaine vaste et il reste encore beaucoup à découvrir concernant le répertoire des capacités cognitives chez les oiseaux domestiques, leur ontogenèse*, leur manière de mettre à profit ces capacités pour s’adapter à leur environnement social ou physique, entre autres. Ces connaissances sont essentielles pour mieux comprendre la biologie de ces oiseaux, leur univers mental et proposer des systèmes d’élevage adaptés aux différents profils d’animaux présents au sein d’un même groupe. Cela pourrait aider au développement de conditions d’élevage bénéfiques au bien-être aussi bien mental que physique des animaux. En retour, certaines recherches s’intéressent à la manière dont les différentes conditions de vie des animaux impactent le développement et la mobilisation de leurs capacités cognitives. Par exemple, quelles sont les conséquences de l’absence de figure de référence lors de la phase sensible sur le développement du poussin ? Ou encore, est-ce que la taille de la cage ou le nombre d’animaux au sol a une influence sur le développement de certaines capacités cognitives chez le jeune adulte, et lesquelles ? Ce type de questionnement a un intérêt particulier dans la compréhension du comportement et du bien-être de ces animaux tout au long du processus d’élevage.
Ces connaissances sur les capacités cognitives des volailles permettront d'envisager la mise en place de nouvelles pratiques pour améliorer le bien-être des animaux, comme les enrichissements cognitifs ou occupationnels. Davantage basés sur les capacités cognitives, ces nouveaux enrichissements pourraient constituer des leviers efficaces pour stimuler les animaux. Proposer un enrichissement dans l'environnement d'élevage est bénéfique au développement des capacités cognitives chez le jeune et permet de diminuer les comportements anormaux. Particulièrement, les enrichissements cognitifs consistent à présenter des challenges aux animaux dans leur milieu de vie : l'animal porte son attention sur un stimulus qui indique un évènement à venir (par exemple un son qui précède toujours la distribution alimentaire), ou encore sur une tâche à résoudre plus ou moins simple (par exemple la mise à disposition d'un dispositif en accès libre, comme un test de choix d'images sur écran, qui permet de gagner une récompense) (Clark, 2017 ; exemple récent chez la poule : Schmelz & Krause, 2021). L'efficacité des enrichissements cognitifs a été montrée chez différentes espèces en réduisant les indicateurs de stress ainsi que certains comportements anormaux (exemple : Puppe et al., 2007 ; Zebunke et al., 2013). De plus, contrairement aux traditionnels enrichissements physiques (i.e. amélioration de l'environnement grâce à la présence d'objets ou de structures adaptées), les enrichissements cognitifs semblent être moins sujets au phénomène d'habituation, c'est-à-dire à une perte d'intérêt. Ils pourraient donc contribuer de manière plus efficace et prolongée à leur bien-être, par exemple en diminuant l'occurrence des comportements anormaux comme le picage, améliorant la qualité de vie des animaux. Des études, qui relieraient enrichissement occupationnel et qualité de vie des volailles en élevage, restent à effectuer afin de comprendre de quelle manière la sollicitation des capacités cognitives des animaux pourra participer à leur bien-être.
Annexe 1 : Glossaire
Apprentissage associatif : Apprentissage qui nait de l’association entre deux ou plusieurs évènements (exemple : apprendre qu’un certain comportement engendre une certaine conséquence). Les conditionnements sont un type d’apprentissage associatif.
Apprentissage / Apprentissage social : L’apprentissage est un processus cognitif qui permet à l’animal d’acquérir une nouvelle information, à court terme ou à long terme. L’apprentissage social concerne le cas particulier des apprentissages qui résultent de l’observation directe d’une action ou d’une réaction d’un congénère.
Attente : L’attente est un processus cognitif lié à l’anticipation d’un évènement, positif ou négatif, qui se traduit par une réponse physiologique (battements cardiaques par exemple) et comportementale (vocalisations par exemple). Pour illustration, lorsque la distribution de nourriture est à heure fixe, l’attente de la distribution peut donner lieu à des comportements d’excitation comme une activité locomotrice plus élevée, et pourra donner lieu à une frustration dans le cas où l’attente n’est pas satisfaite.
Attention : L’attention au sens large est la capacité de l’individu à se concentrer sur un stimulus donné. Elle peut être par exemple mesurée via le temps de regard envers le stimulus en question.
Capacités cognitives : Ensemble des processus mentaux qui permettent à l’animal de recevoir, mémoriser, traiter et analyser les informations de son environnement. Exemples : capacités d’attention, de mémorisation, d’apprentissage, résolution de problèmes, prise de décision.
Catégoriser : Capacité à placer un ensemble d’éléments dans une catégorie ou une autre, en fonction de caractéristiques qu’ils partagent avec les éléments de la catégorie en question. Par exemple, la chaise et le fauteuil appartiennent à la catégorie « meubles ».
Cognition : Désigne l’ensemble des capacités cognitives d’un individu, ainsi que les structures et activités cérébrales qui sont le support de ces capacités cognitives.
Concept : Dans le texte, représentation mentale des caractéristiques qui définissent une catégorie.
Conditionnement : Le conditionnement est un type d’apprentissage associatif. Il existe différents types de conditionnements. Le conditionnement classique consiste à associer un stimulus conditionnel (par exemple un son de cloche) à une réaction inconditionnelle (salivation chez le chien) par l’intermédiaire d’un stimulus inconditionnel (le son de cloche annonce le morceau de viande et déclenche donc la réponse de salivation). Le conditionnement opérant consiste à favoriser – ou diminuer – la survenue d’un comportement de l’animal (par exemple se mettre assis) à la suite d’un indice externe (l’ordre « assis ») en le renforçant positivement (friandise) – ou négativement.
Conditions contrôlées : Une expérience scientifique qui se déroule en conditions contrôlées implique que tout ou majorité des facteurs environnementaux peuvent être contrôlés lors du test, et/ou peuvent être modifiés indépendamment, en contrôlant leur valeur (par exemple le niveau d’éclairage).
Conditions expérimentales : Conditions dans lesquelles l’expérience est réalisée, notamment environnement de test, dispositif de test, fonctionnement et déroulé du test.
Contagion émotionnelle : Transfert rapide d’une émotion d’un individu-sujet (en train de ressentir l’émotion) à un individu-observateur. Ce transfert se fait au travers d’indices comportementaux et/ou physiologiques perçus par l’individu-observateur, qui ressent à son tour l’émotion sans en avoir été le sujet direct.
Émotion : État affectif ponctuel en réaction à un évènement. Cet état correspond à une expérience mentale subjective, et est accompagnée par des signes physiologiques (variation de température, battements de cœur, respiration, variations de taux de certains métabolites).
Empathie : Capacité à se mettre à la place de l’autre, comprendre son point de vue et/ou identifier ses émotions. L’empathie est un type de contagion émotionnelle mais elle implique des processus cognitifs plus élaborés.
Empreinte : Processus d’attachement social chez l’oiseau qui a lieu peu après l’éclosion, lors de la période sensible, et qui est ordinairement dirigé vers la mère.
Flexibilité comportementale : Capacité à ajuster son comportement plus ou moins rapidement face à des situations changeantes.
Fourragement : Recherche de nourriture par exploration de l’environnement.
Individu démonstrateur : L’individu démonstrateur est celui qui a appris, avant le test, un comportement particulier (par exemple : préférer un type de nourriture ou en rejeter un autre). Lors du test, un individu observateur/naïf est mis en présence de l’individu démonstrateur. Généralement, l’analyse s’intéresse au comportement de l’individu naïf pendant ou après avoir été confronté au comportement de l’individu démonstrateur.
Interaction sociale : Échange dirigé au cours duquel une information est partagée entre deux ou plusieurs individus. Exemples : expressions agressives de dominance, toilettage mutuel, communication.
Latéralisation : Développement différencié entre l’hémisphère droit de l’encéphale (« cerveau droit ») et l’hémisphère gauche (« cerveau gauche »). Cette latéralisation, qui peut être importante chez les oiseaux, peut engendrer des différences de traitement cognitif des informations si celles-ci sont perçues uniquement par l’œil droit ou l’œil gauche par exemple.
Mémoire déclarative : Mémoire des faits et des évènements. Elle est caractérisée par une mémorisation à long terme et par son aspect explicite, c’est-à-dire que l’individu est capable d’exprimer, par un comportement, les informations qu’il a en mémoire (exemple des souvenirs).
Mémorisation : Capacité à enregistrer et conserver des informations venant d'évènements divers. Il existe différentes formes de mémorisation à court ou long terme, conscientes ou inconscientes.
Métamémoire : Ensemble des connaissances qu’un individu a sur le contenu de sa mémoire, et qui lui permet d’évaluer mentalement le niveau de ses connaissances. Autrement dit, la métamémoire est la capacité à savoir si l’on a une certaine information en mémoire (si l’on « sait ») ou non.
Neurotransmission : Processus de transmission de l’information nerveuse entre neurones.
Ontogénèse : Processus de développement physiologique et psychologique d’un individu.
Perception : Dans notre utilisation du terme, la perception d’une situation par l’animal reflète son ressenti (positif, négatif ou neutre) d’une situation donnée. Elle dépend donc de l’information à laquelle il a accès avec ses sens, mais aussi de la manière dont il est capable de comprendre la situation grâce à ses capacités cognitives.
Période sensible : Chez l’oiseau, la période sensible est la période au cours de laquelle le processus d’attachement social (empreinte) a lieu, à travers l’attachement à la mère. Cette période sensible se déroule rapidement dans les premiers jours de vie de l’animal.
Personnalité : Combinaison de traits comportementaux propre à chaque individu (exemple : sociabilité, témérité), qui reste constante quel que soit le contexte et tout au long de la vie de l’animal. La personnalité se détermine au cours du développement de l’animal, en fonction de sa génétique et de son environnement de vie.
Picage sévère : Comportement de picage dont la fréquence et l’intensité sont anormalement exacerbés, et qui se traduit par la présence de lésions corporelles et la perte de plumes (picage agressif ou picage des plumes) portant ainsi atteinte au bien-être des animaux. Il est à distinguer du picage au sens strict, qui est un comportement naturel d’interactions sociales affiliatives (picage doux) ou agressives (picage agressif).
Processus mentaux : Parmi les capacités cognitives, les processus mentaux (ou processus cognitifs) désignent les différentes fonctions de traitement de l’information, c’est-à-dire les systèmes par lesquels l’information est analysée et intégrée dans les connaissances.
Raisonnement : Processus mental composite qui permet d’analyser une situation, et éventuellement en tirer une conclusion, par le biais d’inférences (connaissance des liens entre différents éléments) plus ou moins complexes ou abstraites.
Self-contrôle moteur : Capacité d’inhibition comportementale, qui renseigne sur la capacité d’un individu à refreiner son impulsivité motrice (ne pas déclencher ou arrêter un mouvement, un déplacement).
Sociabilité : Trait de personnalité qui qualifie la tendance relative de l’animal à rechercher la compagnie de ses congénères.
Spécialisations adaptatives : Caractéristiques physiologiques ou psychologiques d’une espèce qui sont la conséquence de son histoire évolutive, et donc particulièrement adaptées à son mode de vie naturel.
Statut hiérarchique : Position de l’individu dans la hiérarchie de dominance, au sein du groupe social. Il existe différents systèmes de hiérarchies sociales, comme la hiérarchie linéaire (A→B→C) ou la hiérarchie triangulaire (A→B→C→A) comme chez la poule domestique par exemple.
Théorie de l’esprit : Capacité à se représenter les états mentaux d’un autre individu (ce qu’il pense), en les inférant à partir de l’observation de ses comportements.
Valence : Qualité d’un évènement, aspect émotionnel qui est ressenti, positif ou négatif.
Notes
- Cet article est issu d’une synthèse présentée aux 14èmes Journées de la Recherche Avicole des 9-10 mars 2022 à Tours. (Calandreau et al., 2022)
- Les mots suivis d’un astérisque sont définis dans le Glossaire en Annexe 1.
Références
- Akins, C. K., & Zentall, T. R. (1998). Imitation in Japanese quail: The role of reinforcement of demonstrator responding. Psychonomic Bulletin & Review, 5(4), 694‑697. doi:10.3758/BF03208847
- ANSES (2018). Avis de l’Agence nationale de sécurité sanitaire de l’alimentation, de l’environnement et du travail relatif au « Bien-être animal : Contexte, définition et évaluation ». Saisine n° 2016-SA-0288. https://www.anses.fr/fr/system/files/SABA2016SA0288.pdf
- Bertin, A., Calandreau, L., Arnould, C., Nowak, R., Lévy, F., Noirot, V., Bouvarel, I., & Leterrier, C. (2010). In ovo olfactory experience influences post-hatch feeding behaviour in young chickens. Ethology, 116(11), 1027‑1037. doi:10.1111/j.1439-0310.2010.01820.x
- Bestman, M., Verwer, C., Brenninkmeyer, C., Willett, A., Hinrichsen, L., Smajlhodzic, F., Heerkens, J., Gunnarsson, S., & Ferrante, V. (2017). Feather-pecking and injurious pecking in organic laying hens in 107 flocks from eight European countries. Animal Welfare, 26(3), 355‑363. doi:10.7120/09627286.26.3.355
- Bestman, M., Verwer, C., van Niekerk, T., Leenstra, F., Reuvekamp, B., Amsler-Kepalaite, Z., & Maurer, V. (2019). Factors related to free-range use in commercial laying hens. Applied Animal Behaviour Science, 214, 57‑63. doi:10.1016/j.applanim.2019.02.015
- Bradshaw, R. H., & Dawkins, M. S. (1993). Slides of conspecifics as representatives of real animals in laying hens (Gallus domesticus). Behavioural Processes, 28(3), 165‑172. doi:10.1016/0376-6357(93)90089-A
- Calandreau L., Ferreira V. H. B., Degrande R., & Guesdon V. (2022). Comment les volailles perçoivent-elles leur environnement ? Quatorzièmes Journées de la Recherche Avicole et Palmipèdes à Foie Gras, Tours. 13(5), 678-679. doi:10.1016/j.anscip.2022.05.144
- Campbell, D. L. M., Dickson, E. J., & Lee, C. (2019). Application of open field, tonic immobility, and attention bias tests to hens with different ranging patterns. PeerJ, 7. doi:10.7717/peerj.8122
- Campbell, D. L. M., Dyall, T. R., Downing, J. A., Cohen-Barnhouse, A. M., & Lee, C. (2020). Rearing enrichments affected ranging behavior in free-range laying hens. Frontiers in Veterinary Science, 7. https://www.frontiersin.org/article/10.3389/fvets.2020.00446
- Campbell, D., Bari, M. S., & Rault, J.-L. (2021). Free-range egg production: its implications for hen welfare. Animal Production Science, 61(10), 848-855. doi:10.1071/AN19576
- Clark, F. (2017). Cognitive enrichment and welfare: Current Approaches and future directions. Animal Behavior and Cognition, 4(1), 52‑71. doi:10.12966/abc.05.02.2017
- Cloutier, S., Newberry, R., Honda, K., & Alldredge, J. R. (2002). Cannibalistic behaviour spread by social learning. Animal Behaviour, 63(6), 1153‑1162. doi:10.1006/anbe.2002.3017
- Coppens, C. M., de Boer, S. F., & Koolhaas, J. M. (2010). Coping styles and behavioural flexibility: towards underlying mechanisms. Philosophical Transactions of the Royal Society B: Biological Sciences, 365(1560), 4021‑4028. doi:10.1098/rstb.2010.0217
- Coton, J., Guinebretière, M., Guesdon, V., Chiron, G., Mindus, C., Laravoire, A., Pauthier, G., Balaine, L., Descamps, M., Bignon, L., Huneau-Salaün, A., & Michel, V. (2019). Feather pecking in laying hens housed in free-range or furnished-cage systems on French farms. British Poultry Science, 60(6), 617‑627. doi:10.1080/00071668.2019.1639137
- Cozzutti, C., & Vallortigara, G. (2001). Hemispheric memories for the content and position of food caches in the domestic chick. Behavioral Neuroscience, 115(2), 305‑313. doi:10.1037/0735-7044.115.2.305
- Cronin, G. M., Glatz, P. C., Cronin, G. M., & Glatz, P. C. (2020). Causes of feather pecking and subsequent welfare issues for the laying hen: a review. Animal Production Science, 61(10), 990‑1005. doi:10.1071/AN19628
- Daisley, J. N., Vallortigara, G., & Regolin, L. (2010). Logic in an asymmetrical (social) brain: transitive inference in the young domestic chick. Social Neuroscience, 5(3), 309‑319. doi:10.1080/17470910903529795
- Dawkins, M. S., Cook, P. A., Whittingham, M. J., Mansell, K. A., & Harper, A. E. (2003). What makes free-range broiler chickens range? In situ measurement of habitat preference. Animal Behaviour, 66(1), 151‑160. doi:10.1006/anbe.2003.2172
- Degrande, R., Cornilleau, F., Lansade, L., Jardat, P., Colson, V., & Calandreau, L. (2022). Domestic hens succeed at serial reversal learning and perceptual concept generalisation using a new automated touchscreen device. Animal, 16(8). doi:10.1016/j.animal.2022.100607
- DeMello, L. R., Foster, T. M., & Temple, W. (1993). The effect of increased response requirements on discriminative performance of the domestic hen in a visual acuity task. Journal of the Experimental Analysis of Behavior, 60(3), 595-609. doi:10.1901/jeab.1993.60-595
- Dougherty, L., & Guillette, L. (2018). Linking personality and cognition: a meta-analysis. Philosophical Transactions of the Royal Society B: Biological Sciences, 373. doi:10.1098/rstb.2017.0282
- Edgar, J. L., Lowe, J. C., Paul, E. S., & Nicol, C. J. (2011). Avian maternal response to chick distress. Proceedings of the Royal Society B: Biological Sciences, 278(1721), 3129‑3134. doi:10.1098/rspb.2010.2701
- Edgar, J. L., Paul, E. S., & Nicol, C. J. (2013). Protective mother hens: cognitive influences on the avian maternal response. Animal Behaviour, 86(2), 223‑229. doi:10.1016/j.anbehav.2013.05.004
- FAO (2010). Systèmes d’élevage ; Répartition mondiale. Consulté le 19 juillet 2022 sur. https://www.fao.org/livestock-systems/global-distributions/fr/
- Ferreira, V. H. B., Peuteman, B., Lormant, F., Valenchon, M., Germain, K., Brachet, M., Leterrier, C., Lansade, L., Calandreau, L., & Guesdon, V. (2019). Relationship between ranging behavior and spatial memory of free-range chickens. Behavioural Processes, 166. doi:10.1016/j.beproc.2019.103888
- Ferreira, V. H. B., Barbarat, M., Lormant, F., Germain, K., Brachet, M., Løvlie, H., Calandreau, L., & Guesdon, V. (2020a). Social motivation and the use of distal, but not local, featural cues are related to ranging behavior in free-range chickens (Gallus gallus domesticus). Animal Cognition, 23(4), 769‑780. doi:10.1007/s10071-020-01389-w
- Ferreira, V. H. B., Reiter, L., Germain, K., Calandreau, L., & Guesdon, V. (2020b). Uninhibited chickens: ranging behaviour impacts motor self-regulation in free-range broiler chickens (Gallus gallus domesticus). Biology Letters, 16(1). doi:10.1098/rsbl.2019.0721
- Ferreira, V. H. B., Guesdon, V., & Calandreau, L. (2021). How can the research on chicken cognition improve chicken welfare: a perspective review. World’s Poultry Science Journal, 77(3), 679-698. doi:10.1080/00439339.2021.1924920
- Ferreira, V. H. B., Simoni, A., Germain, K., Leterrier, C., Lansade, L., Collin, A., Mignon-Grasteau, S., Le Bihan-Duval, E., Guettier, E., Leruste, H., Løvlie, H., Calandreau, L., & Guesdon, V. (2022). Foraging behavior shows individual-consistency over time, and predicts range use in slow-growing free-range male broiler chickens. Frontiers in Veterinary Science, 9. https://www.frontiersin.org/articles/10.3389/fvets.2022.814054
- Forkman, B. (2000). Domestic hens have declarative representations. Animal Cognition, 3(3), 135‑137. doi:10.1007/s100710000074
- Freire, R. (2020). Understanding chicken learning and cognition and implications for improved management. In Burleigh Dodds series in agricultural science (p. 91‑116). Burleigh Dodds Science Publishing. doi:10.19103/AS.2020.0078.04
- Freire, R., & Hazel, S. J. (2017). Are chicken minds special? Animal Sentience, 17(4). doi:10.51291/2377-7478.1223
- Freire, R., Munro, U. H., Rogers, L. J., Wiltschko, R., & Wiltschko, W. (2005). Chickens orient using a magnetic compass. Current Biology, 15(16), R620-621. doi:10.1016/j.cub.2005.08.017
- Fritz, J., Bisenberger, A., & Kotrschal, K. (2000). Stimulus enhancement in greylag geese: socially mediated learning of an operant task. Animal Behaviour, 59(6), 1119‑1125. doi:10.1006/anbe.2000.1424
- Garnham, L., & Løvlie, H. (2018). Sophisticated fowl: the complex behaviour and cognitive skills of chickens and red junglefowl. Behavioral Sciences, 8(1), 13. doi:10.3390/bs8010013
- Gebhardt-Henrich, S. G., Toscano, M. J., & Fröhlich, E. K. F. (2014). Use of outdoor ranges by laying hens in different sized flocks. Applied Animal Behaviour Science, 155, 74‑81. doi:10.1016/j.applanim.2014.03.010
- Harlander-Matauschek, A., Wassermann, F., Zentek, J., & Bessei, W. (2008). Laying Hens Learn to Avoid Feathers. Poultry science, 87(9), 1720‑1724. doi:10.3382/ps.2007-00510
- Harlander-Matauschek, A., Beck, P., & Rodenburg, T. B. (2010). Effect of an early bitter taste experience on subsequent feather-pecking behaviour in laying hens. Applied Animal Behaviour Science, 127(3‑4), 108‑114. doi:10.1016/j.applanim.2010.09.005
- Heinsius, J., Van Staaveren, N., Kwon, I. Y., Li, A., Kjaer, J. B., & Harlander-Matauschek, A. (2020). Chickens selected for feather pecking can inhibit prepotent motor responses in a Go/No-Go task. Scientific Reports, 10, 6485. doi:10.1038/s41598-020-63618-z
- Hogue, M.-E., Beaugrand, J. P., & Laguë, P. C. (1996). Coherent use of information by hens observing their former dominant defeating or being defeated by a stranger. Behavioural Processes, 38(3), 241‑252. doi:10.1016/S0376-6357(96)00035-6
- Iwaniuk, A., & Wylie, D. (2020). Sensory systems in birds: what we have learned from studying sensory specialists. Journal of Comparative Neurology, 528(17), 2902‑2918. doi:10.1002/cne.24896
- Jung, L., & Knierim, U. (2018). Are practice recommendations for the prevention of feather pecking in laying hens in non-cage systems in line with the results of experimental and epidemiological studies? Applied Animal Behaviour Science, 200, 1‑12. doi:10.1016/j.applanim.2017.10.005
- Karakashian, S. J., Gyger, M., & Marler, P. (1988). Audience effects on alarm calling in chickens (Gallus gallus). Journal of Comparative Psychology, 102(2), 129‑135. doi:10.1037/0735-7036.102.2.129
- Kjaer, J. B. (2009). Feather pecking in domestic fowl is genetically related to locomotor activity levels: implications for a hyperactivity disorder model of feather pecking. Behavior Genetics, 39(5), 564‑570. doi:10.1007/s10519-009-9280-1
- Larsen, H., Hemsworth, P., Cronin, G., Gebhardt-Henrich, S., Smith, K., & Rault, J.-L. (2018). Relationship between welfare and individual ranging behaviour in commercial free-range laying hens. Animal, 12(11), 2356-2364. doi:10.1017/S1751731118000022
- Le Neindre, P., Bernard, E., Boissy, A., Boivin, X., Calandreau, L., Delon, N., Deputte, B., Desmoulin‐Canselier, S., Dunier, M., Faivre, N., Giurfa, M., Guichet, J., Lansade, L., Larrère, R., Mormède, P., Prunet, P., Schaal, B., Servière, J., & Terlouw, C. (2017). Animal Consciousness. EFSA Supporting Publications, 14(4). doi:10.2903/sp.efsa.2017.EN-1196
- Lea, S. E. G., Slater, A. M., & Ryan, C. M. E. (1996). Perception of object unity in chicks: a comparison with the human infant. Infant Behavior and Development, 19(4), 501‑504. doi:10.1016/S0163-6383(96)90010-7
- Loconsole, M., Mascalzoni, E., Daisley, J. N., De Agrò, M., Vallortigara, G., & Regolin, L. (2021). Lateralized declarative-like memory for conditional spatial information in domestic chicks (Gallus gallus). Symmetry, 13(5), 906. doi:10.3390/sym13050906
- Lormant, F., Cornilleau, F., Constantin, P., Meurisse, M., Lansade, L., Leterrier, C., Lévy, F., & Calandreau, L. (2020). Research note: role of the hippocampus in spatial memory in Japanese quail. Poultry Science, 99(1), 61‑66. doi:10.3382/ps/pez507
- Marino, L. (2017). Thinking chicken: a review of cognition, emotion, and behavior in the domestic chicken. Animal Cognition, 20(2), 127‑147. doi:10.1007/s10071-016-1064-4
- Marler, P., Pickert, R., & Gyger, M. (1987). Semantics of an avian alarm call system: the male domestic fowl, Gallus domesticus. Behaviour, 102(1‑2), 15‑39. doi:10.1163/156853986X00027
- Martin, G. M., & Bellingham, W. P. (1979). Learning of visual food aversions by chickens (Gallus gallus) over long delays. Behavioral and Neural Biology, 25(1), 58‑68. doi:10.1016/S0163-1047(79)90756-8
- McCabe, B. J. (2019). Visual imprinting in birds: behavior, models, and neural mechanisms. Frontiers in Physiology, 10. doi:10.3389/fphys.2019.00658
- Nakagawa, S., Etheredge, R. J. M., Foster, T. M., Sumpter, C. E., & Temple, W. (2004). The effects of changes in consequences on hens’ performance in delayed-matching-to-sample tasks. Behavioural Processes, 67(3), 441‑451. doi:10.1016/j.beproc.2004.07.005
- Nakamori, T., Maekawa, F., Sato, K., Tanaka, K., & Ohki-Hamazaki, H. (2013). Neural basis of imprinting behavior in chicks. Development, Growth & Differentiation, 55(1), 198‑206. doi:10.1111/dgd.12028
- Nakamura, N., Watanabe, S., Betsuyaku, T., & Fujita, K. (2011). Do birds (pigeons and bantams) know how confident they are of their perceptual decisions? Animal Cognition, 14(1), 83‑93. doi:10.1007/s10071-010-0345-6
- Newberry, R. C., Keeling, L. J., Estevez, I., & Bilčík, B. (2007). Behaviour when young as a predictor of severe feather pecking in adult laying hens: The redirected foraging hypothesis revisited. Applied Animal Behaviour Science, 107(3-4), 262-274. doi:10.1016/j.applanim.2006.10.010
- Nicol, C. J. (2004). Chicken Cognition. Dans Welfare of the Laying Hen. « 27th Poultry Science Symposium of the World's Poultry Science Association (UK Branch) » (Bristol, UK, July 2003), 177‑189, CABI Publishing. https://www.cabidigitallibrary.org/doi/10.1079/9780851998138.0177
- Nicol, C. J. (2015). The behavioural biology of chickens. CABI. https://www.cabidigitallibrary.org/doi/book/10.1079/9781780642499.0000
- Nordquist, R. E., Heerkens, J. L. T., Rodenburg, T. B., Boks, S., Ellen, E. D., & van der Staay, F. J. (2011). Laying hens selected for low mortality: behaviour in tests of fearfulness, anxiety and cognition. Applied Animal Behaviour Science, 131(3‑4), 110‑122. doi:10.1016/j.applanim.2011.02.008
- Pettersson, I. C., Freire, R., & Nicol, C. J. (2016). Factors affecting ranging behaviour in commercial free-range hens. World’s Poultry Science Journal, 72(1), 137‑150. doi:10.1017/S0043933915002664
- Pichová, K., Košťál, Ľ., De Haan, T. I., Van der Eijk, J. A., & Rodenburg, T. (2021). High and low feather pecking selection lines of laying hens differ in response to a judgment bias test. Applied Animal Behaviour Science, 238. doi:10.1016/j.applanim.2021.105305
- Puppe, B., Ernst, K., Schön, P. C., & Manteuffel, G. (2007). Cognitive enrichment affects behavioural reactivity in domestic pigs. Applied Animal Behaviour Science, 105(1), 75‑86. doi:10.1016/j.applanim.2006.05.016
- Regolin, L., & Vallortigara, G. (1995). Perception of partly occluded objects by young chicks. Perception & Psychophysics, 57(7), 971‑976. doi:10.3758/BF03205456
- Regolin, L., Rugani, R., Stancher, G., & Vallortigara, G. (2011). Spontaneous discrimination of possible and impossible objects by newly hatched chicks. Biology Letters, 7(5), 654‑657. doi:10.1098/rsbl.2011.0051
- Rodenburg, T. B., Van Hierden, Y. M., Buitenhuis, A. J., Riedstra, B., Koene, P., Korte, S. M., Van der Poel, J. J., Groothuis, T. G. G., & Blokhuis, H. J. (2004). Feather pecking in laying hens: new insights and directions for research? Applied Animal Behaviour Science, 86(3‑4), 291‑298. doi:10.1016/j.applanim.2004.02.007
- Rugani, R., Fontanari, L., Simoni, E., Regolin, L., & Vallortigara, G. (2009). Arithmetic in newborn chicks. Proceedings of the Royal Society B: Biological Sciences, 276(1666), 2451‑2460. doi:10.1098/rspb.2009.0044
- Rugani, R., Vallortigara, G., Priftis, K., & Regolin, L. (2015). Number-space mapping in the newborn chick resembles humans’ mental number line. Science, 347(6221), 534‑536. doi:10.1126/science.aaa1379
- Salva, O. R., Daisley, J. N., Regolin, L., & Vallortigara, G. (2009). Lateralization of social learning in the domestic chick, Gallus gallus domesticus: learning to avoid. Animal Behaviour, 78(4), 847‑856. doi:10.1016/j.anbehav.2009.06.021
- Schmelz, M., & Krause, E. T. (2021). Simple but complex—a laying hen study as proof of concept of a novel method for cognitive enrichment and research. Frontiers in Animal Science, 2. doi:10.3389/fanim.2021.671905
- Sih, A., & Del Giudice, M. (2012). Linking behavioural syndromes and cognition: a behavioural ecology perspective. Philosophical Transactions of the Royal Society B: Biological Sciences, 367(1603), 2762‑2772. doi:10.1098/rstb.2012.0216
- Taylor, P., Hemsworth, P., Groves, P., Gebhardt-Henrich, S., & Rault, J.-L. (2017a). Ranging behaviour of commercial free-range broiler chickens 1: Factors related to flock variability. Animals, 7(7). doi:10.3390/ani7070054
- Taylor, P., Hemsworth, P., Groves, P., Gebhardt-Henrich, S., & Rault, J.-L. (2017b). Ranging behaviour of commercial free-range broiler chickens 2: Individual variation. Animals 7(7). doi:10.3390/ani7070055
- Vallortigara, G., Zanforlin, M., & Pasti, G. (1990). Geometric modules in animals’ spatial representations: a test with chicks (Gallus gallus domesticus). Journal of Comparative Psychology, 104(3), 248‑254. doi:10.1037/0735-7036.104.3.248
- Van Staaveren, N., & Harlander, A. (2020). Cause and prevention of injurious pecking in chickens. Burleigh Dodds Science Publishing. doi:10.19103/AS.2020.0078.15
- Vauclair, J., & Kreutzer, M. (2004). L’éthologie cognitive. Ophrys - Maison des sciences de l’homme. doi:10.4000/books.editionsmsh.7081
- Weiß, B. M., Kehmeier, S., & Schloegl, C. (2010). Transitive inference in free-living greylag geese, Anser anser. Animal Behaviour, 79(6), 1277‑1283. doi:10.1016/j.anbehav.2010.02.029
- Wiltschko, R., Nießner, C., & Wiltschko, W. (2021). The magnetic compass of birds: the role of cryptochrome. Frontiers in Physiology, 12. doi:10.3389/fphys.2021.667000
- Zanforlin, M. (1981). Visual perception of complex forms (anomalous surfaces) in chicks. Italian Journal of Psychology, 8(1), 1‑16.
- Zebunke, M., Puppe, B., & Langbein, J. (2013). Effects of cognitive enrichment on behavioural and physiological reactions of pigs. Physiology & Behavior, 118, 70‑79. doi:10.1016/j.physbeh.2013.05.005
- Zeltner, E., Klein, T., & Huber-Eicher, B. (2000). Is there social transmission of feather pecking in groups of laying hen chicks? Animal Behaviour, 60(2), 211216. doi:10.1006/anbe.2000.1453
- Zidar, J., Balogh, A., Favati, A., Jensen, P., Leimar, O., Sorato, E., & Løvlie, H. (2018). The relationship between learning speed and personality is age- and task-dependent in red junglefowl. Behavioral Ecology and Sociobiology, 72(10), 168. doi:10.1007/s00265-018-2579-2
- Zimmerman, P. H., Buijs, S. A. F., Bolhuis, J. E., & Keeling, L. J. (2011). Behaviour of domestic fowl in anticipation of positive and negative stimuli. Animal Behaviour, 81(3), 569‑577. doi:10.1016/j.anbehav.2010.11.028
Résumé
Les capacités cognitives rassemblent l’ensemble des processus mentaux comme l’attention, la mémorisation, l’apprentissage ou encore les capacités de raisonnement. Ces capacités cognitives permettent à un animal de comprendre et s’adapter à son environnement et sont à la base de très nombreux comportements. Dans cette revue, nous proposons dans une première partie un état des lieux des connaissances actuelles sur les capacités cognitives des volailles, en nous concentrant sur la poule pondeuse et le poulet de chair qui sont les plus étudiés en cognition. Nous montrons brièvement que ces oiseaux possèdent des sens diversifiés et développés, qu’ils interagissent avec leurs congénères et qu’ils sont capables d’apprentissages, entre autres. Même s’il reste encore des recherches à mener, ces oiseaux possèdent un répertoire de capacités cognitives bien plus étendu que ce qui est souvent imaginé. Dans une seconde partie, nous illustrons en quoi ces connaissances apportent une vision nouvelle de notre compréhension de comportements observés en élevage, comme l’exploration du parcours chez le poulet élevé en plein air, ou le picage chez les poules pondeuses. La thématique d’étude de la cognition animale, qui se développe de façon croissante au niveau international, contribue à mieux comprendre la biologie de ces animaux. Sur le long terme, les recherches en cognition permettront de proposer des pistes originales pour guider les pratiques d’élevage et favoriser le bien-être des animaux d’élevage.
Pièces jointes
Pas de document complémentaire pour cet articleStatistiques de l'article
 Vues: 6329
Vues: 6329
Téléchargements
 PDF: 593
PDF: 593
 XML: 117
XML: 117
Articles les plus lus par le même auteur ou la même autrice
- Frédérique PITEL, Fanny CALENGE, Nadège AIGUEPERSE, Jordi ESTELLÉ-FABRELLAS, Vincent COUSTHAM, Ludovic CALANDREAU, Mireille MORISSON, Pascale CHAVATTE-PALMER, Cécile GINANE, Rôle de l'environnement précoce dans la variabilité des phénotypes et l’adaptation des animaux d’élevage à leur milieu , INRAE Productions Animales: Vol. 32 No 2 (2019): Volume 32 Numéro 2 : Numéro spécial. De grands défis et des solutions pour l'élevage
- Karine GERMAIN, Claire BONNEFOUS, Ludovic CALANDREAU, Geoffrey CHIRON, Brieuc DESAINT, Elisabeth LE BIHAN-DUVAL, Vitor FERREIRA, Vanessa GUESDON, Laurence A. GUILLOTEAU, Sarah LOMBARD, Bertrand MÉDA, Anne SILVESTRE, Anne COLLIN, L’élevage de poulets biologiques : les verrous à lever et les opportunités offertes par cette production , INRAE Productions Animales: Vol. 37 No 2 (2024): Numéro spécial : L’élevage biologique : conditions et potentiel de développement