

Santé et bien-être de la truie gestante et du porc en croissance
Chapeau
Les maladies dites « de production » sont des maladies multifactorielles, infectieuses ou non, qui affectent la santé et le bien-être des animaux et limitent leur productivité et celle des élevages. Elles ont également été définies comme persistant et augmentant avec l’intensification de l’élevage. Pour apporter des solutions pour prévenir ces maladies et/ou en atténuer les effets chez les porcs et les volailles, un programme européen PROHEALTH (FP7 n°613574 ; 2013 - 2018) a été réalisé. Les résultats d’INRAE dans ce projet sont présentés ici
Introduction
En Europe, la grande majorité des porcs et des poulets de chair sont élevés dans des élevages intensifs, en bâtiments fermés avec une forte densité d’animaux. Contrairement aux idées reçues, les maladies de production, telles que définies dans le chapeau, ne sont pas spécifiques de ce type d’élevage soumis à forte contrainte de productivité, mais dans ces derniers, certaines pratiques peuvent constituer des facteurs aggravants de ces maladies et de leurs conséquences. Pour répondre aux préoccupations citoyennes et sociétales et aux enjeux de santé publique, l’amélioration de la santé des animaux d’élevage doit concilier une meilleure prise en compte de leur bien-être et une réduction de l'utilisation des antibiotiques. L’ambition du projet PROHEALTH était d’apporter des connaissances et de proposer des solutions pour prévenir et réduire les effets des maladies de production des porcs et des volailles. Ce projet interdisciplinaire d’ampleur (figure 1) a rassemblé 22 partenaires, chercheurs en science vétérinaire, épidémiologie, physiologie, génétique, nutrition, sciences humaines et sociales et des acteurs des filières. Cet article synthétise les principaux résultats obtenus par INRAE pour l’espèce porcine.
Figure 1. Les différentes tâches et la liste des partenaires du projet PROHEALTH.

Chaque rectangle (traits pleins) correspond à une tâche. Les flèches illustrent les relations entre tâches. Les unités INRAE participantes et leurs principaux collaborateurs sont indiqués en gris. Les résultats présentés dans cet article sont issus des tâches « Survie néonatale » et « Génétique ».
Les maladies de production affectant les porcs sont diverses, tant par le stade physiologique que par le système biologique concernés. Elles affectent les revenus des éleveurs (tableau 1) en induisant des pertes animales (mortalité, ralentissement de la croissance, réforme), des coûts liés à l'utilisation de médicaments et du travail supplémentaire (PROHEALTH, 2015). Un des objectifs de PROHEALTH était de proposer des indicateurs pour quantifier l'effet de ces maladies et d'évaluer l'efficacité de solutions préventives sur les animaux. En plus des approches expérimentales décrites ci-après, des indicateurs (encadré 1) ont été identifiés par cartographie systémique de la littérature (Stavrakakis et al., 2019). Par ailleurs, différents leviers susceptibles de modifier la sensibilité des porcs aux maladies de production ont été testés expérimentalement. Parmi les leviers étudiés, nous nous sommes intéressés à l'influence des conditions de logement des truies pendant la gestation sur la survie des porcelets, à l'influence de l'aliment sur la santé et les performances des porcelets au sevrage et à celle des conditions de logement des porcs en croissance sur la survenue de troubles locomoteurs, respiratoires et inflammatoires. Par ailleurs, un facteur de risque des maladies de production souvent évoqué lors de la dernière décennie (Prunier et al., 2010 ; Knap et Rauw, 2009) est la sélection génétique sur des critères de productivité mais cela n'est pas clairement établi chez le porc. Les lignées divergentes sélectionnées sur la Consommation Moyenne Journalière Résiduelle (CMJR (ATOL_0002160)
Tableau 1. Le coût des maladies de production chez le porc et la truie (d’après PROHEALTH consortium, 2015).
Maladies respiratoires |
4,2 €/porc |
|---|---|
Mortalité |
12 à 23 €/portée (avant le sevrage) |
Maladies digestives |
3,4 €/porc pour diarrhées post-sevrage |
Boiteries |
145 à 180 €/truie |
Mammite et syndrôme MMA |
95 €/truie |
Parasites |
7 €/porc pour infestation par Ascaris suum |
Cannibalisme (caudophagie) |
2 €/porc |
Le coût par animal atteint a été calculé d’après les résultats d’une revue systématique de la littérature conduite sur 120 publications. Il intègre le coût des médicaments (antimicrobiens et vaccins), le travail supplémentaire, l’impact sur les performances et les pertes en animaux (réforme, mortalité).
Encadré 1. Identification, par cartographie systémique de la littérature et méta-analyse, de variables pouvant être utilisées comme indicateurs de maladies de production (d'après Stavrakakis et al., 2019).
Démarche
Extraction de données issues d’animaux témoins et malades dans 67 articles sélectionnés à partir de 2 339 articles traitant des maladies de production.
Méta-analyse des variables ayant plus de 5 occurrences.
Principaux résultats
Plus de 524 variables permettent de caractériser la réponse des animaux aux maladies. Les variables sont des paramètres mesurés dans des prélèvements biologiques (sang, tissus…) ou à l’échelle de l’animal (performances, comportements…).
Le travail met en évidence un manque de données sur certaines maladies (troubles locomoteurs) ou certaines réponses (comportements).
Le Gain Moyen Quotidien (GMQ), la Consommation Moyenne Journalière (CMJ) et les réponses immunitaires sont les indicateurs les plus souvent mesurés. Ces derniers requièrent un prélèvement de sang et leurs mesures peuvent être compliquées à mettre en œuvre (coût, technicité, temps de présence dans le sang…). 
1. Réduction du stress maternel et survie néonatale
1.1. Influence des conditions d’élevage des truies gestantes sur la survie néonatale
La truie donne naissance à de nombreux jeunes relativement immatures à la naissance. Les nouveau-nés sont en compétition pour l'accès aux mamelles et les moins compétitifs meurent rapidement (Edwards, 2002). La sélection génétique sur la prolificité des truies s'est accompagnée d'une augmentation de la mortalité des porcelets. En France comme dans d'autres pays européens, en moyenne un porcelet sur cinq meurt entre la naissance et le sevrage (source IFIP GT-Porc 2020). Entre 2009 et 2016, même si ce taux de pertes sur nés totaux de 20 % est resté stable en France, le nombre de porcelets qui mouraient a continué à augmenter parallèlement à l'augmentation de la prolificité des truies (+ 0,7 porcelet par portée sur cette période). Depuis 2016, le taux de pertes augmente de nouveau légèrement en France (source IFIP GT-Porc 2020). La mortalité des porcelets persistant dans les élevages intensifs et augmentant avec l'intensité de production, elle entre bien dans la définition des maladies de production (Edwards et al., 2016). Cette mortalité génère des pertes économiques (voir tableau 1) et n'est éthiquement pas acceptable.
Plus des deux tiers de la mortalité avant le sevrage se produisent dans les 72 h suivant la naissance (Quesnel et al., 2015). Cette mortalité néonatale dépend de facteurs maternels (capacité utérine, déroulement de la mise bas, production de colostrum et de lait, transfert d'anticorps et de cellules immunitaires, comportement maternel) et du porcelet (poids, maturité et vigueur à la naissance, acquisition de l'immunité), mais aussi de la portée (taille, poids, hétérogénéité) et de l'environnement (Quesnel et al., 2015). Pour améliorer la survie des porcelets, l'accent a été porté sur les qualités maternelles de la truie (génétique, alimentation) et les soins aux jeunes (injection de fer, lampe chauffante, adoptions…) (Baxter et al., 2013). Les conséquences du stress maternel ont été moins étudiées (Merlot et al., 2015). Pourtant, dans les élevages intensifs, les modes de logement et de conduite pendant la gestation peuvent être néfastes au bien-être des truies. Celles-ci y sont en effet soumises à une restriction alimentaire et spatiale, à un stress social lors de la mise en groupe ou de la compétition au distributeur d'aliment, ainsi qu'à une pauvreté des stimuli sensoriels et cognitifs. Ces facteurs sont susceptibles d'activer les réponses physiologiques de stress (Kranendonk et al., 2008). Or dans nombre d'espèces de mammifères, les situations de stress peuvent conditionner les comportements maternels et influencer le développement fœtal et l'immunité des jeunes, et pourraient ainsi se répercuter sur leur survie (Rutherford et al., 2012 ; Merlot et al., 2015). Dans une étude préliminaire, nous avons montré qu'enrichir le milieu d'élevage des truies pendant la gestation en fournissant une litière de paille profonde et en augmentant la superficie disponible réduisait le niveau de stress des truies (Merlot et al., 2017). Ce milieu enrichi était aussi associé à une moindre mortalité des porcelets.
Dans le cadre du projet PROHEALTH, nous avons cherché à comprendre par quelles voies biologiques l’environnement des truies gestantes influence la survie des porcelets (figure 2). Pour cela, nous avons comparé deux milieux d’élevage contrastés pour les truies gestantes en groupe. Ces deux milieux étaient mis en œuvre au sein de l’élevage expérimental de la Chambre Régionale d’Agriculture de Bretagne (CRAB, Saint-Nicolas du Pélem, France). Le premier milieu est dit « conventionnel » (C) car il est présent dans 95 % des élevages français et aussi très répandu dans la plupart des pays européens. C’était un milieu sur caillebotis avec une superficie allouée de 2,4 m2 par truie pour une bande de 24 truies, soit légèrement plus que les 2,25 m2 exigés par la réglementation européenne. Dans le milieu dit « enrichi » (E), les 24 truies de chaque bande disposaient d’une litière profonde de paille et d’une surface de 3,5 m2 par truie. Une dizaine de jours avant la mise bas, toutes les truies étaient transférées en maternité dans des cases individuelles sur caillebotis, où elles étaient logées et soignées de façon identique pour les deux conditions appliquées en gestation. Dans cette première expérience, nous avons comparé la survie des porcelets et les caractéristiques physiologiques, immunitaires et comportementales des truies et des porcelets selon les conditions d’élevage des truies en gestation. L’étude a été menée en 3 répétitions incluant simultanément les deux milieux.
Figure 2. Hypothèse des voies biologiques par lesquelles l’environnement des truies pendant la gestation influencerait la survie des porcelets.

En bleu, les volets abordés dans deux expériences du projet PROHEALTH.
Les conditions d'élevage contrastées pendant la gestation ont été associées à des taux de mortalité naissance-sevrage différents (25,8 % dans le milieu conventionnel versus 16,7 % dans le système enrichi). La différence se concentrait sur les 72 h qui suivent la naissance (13,6 vs 6,3 %, figure 3A). Dans le milieu conventionnel, les truies pendant la gestation présentaient des concentrations salivaires en cortisol (ATOL_0005349) plus élevées que celles du milieu enrichi, suggérant un niveau de stress significativement plus élevé (figure 3B, Merlot et al., 2019). Cette différence était bien due à l'environnement pendant la gestation puisqu'après leur transfert en maternité, les truies issues des deux milieux présentaient des concentrations en cortisol similaires (figure 3B). Des différences modestes ont été observées sur plusieurs marqueurs sanguins de l'état physiologique des truies en fin de gestation, avant leur transfert en maternité. Les concentrations en haptoglobine (ATOL_0000935), une protéine inflammatoire (Eckersall et Bell, 2010), étaient comparables dans les deux milieux. En revanche, les truies du milieu conventionnel présentaient un nombre de granulocytes neutrophiles et une concentration en hydroperoxydes supérieurs, suggérant un niveau de stimulation du système immunitaire et un niveau de stress oxydatif plus élevés. L'intégrité physique des truies du milieu conventionnel était également dégradée car elles étaient 18 % à présenter des boiteries en fin de gestation, contre 2 % en milieu enrichi (Pastorelli et al., 2016). Cette observation confirme l'impact défavorable des sols durs sur la qualité des aplombs, et que le caillebotis est un facteur de risque dans l'émergence de problèmes locomoteurs (Cador et al., 2014).
Figure 3. Taux de mortalité des porcelets et concentrations salivaires en cortisol des truies en fonction de l’enrichissement du milieu d’élevage pendant la gestation.

Figures 3A et 3B, expérience 1 : milieu conventionnel (C) et milieu sur litière profonde de paille et offrant plus d’espace (enrichi, E ; 109 truies Landrace x Large White et leur portée) ; Figures 3C et 3D, expérience 2 : milieu conventionnel (C), conventionnel enrichi avec des jouets en bois et des granulés de paille (CE) et milieu enrichi (E ; 83 truies Landrace x Large White et leur portée). Les figures 3A et 3C présentent les taux de mortalité des porcelets. La mortalité avant 12 h représente les mort-nés et les morts dans les 12 h après la naissance en pourcentage des nés totaux. Les mortalités 12 - 72 h et 72 h – sevrage sont rapportées au nombre de porcelets nés vivants. Les figures 3B et 3D présentent les concentrations salivaires en cortisol des truies. *** P < 0,001 ; a, b P < 0,05 ; t P < 0,10.
Les conditions d'élevage n'ont influencé ni la durée de mise bas, ni le rythme de naissance des porcelets (ou « chronopart »). De même, le volume de colostrum produit ne différait pas et peu de différences apparaissaient sur sa composition en termes de nutriments, immunoglobulines ou cellules immunitaires. Lors des 24 à 48 h post-partum, les truies du milieu conventionnel tendaient à être plus nerveuses et qu'elles montraient de manière plus marquée une réponse négative à l'approche de l'humain (Pastorelli et al., 2016). Cette réactivité comportementale est susceptible d'accroître la mortalité précoce des porcelets par écrasement (Damm et al., 2005).
Le poids moyen des porcelets à la naissance (ATOL_0000093) ne différait pas significativement entre les deux milieux (1,46 et 1,49 kg), mais la proportion de porcelets légers (< 1,2 kg) était plus élevée dans l'environnement conventionnel (25 vs 20 %). De plus, plusieurs indicateurs physiologiques suggéraient que les porcelets nés des truies du milieu conventionnel présentaient un léger désavantage à la naissance pour leur survie future (Quesnel et al., 2019a). Notamment l'étude effectuée sur un sous-effectif de porcelets a montré que les nouveau-nés du milieu conventionnel avaient un tractus gastro-intestinal moins développé et moins de glycogène musculaire, deux caractéristiques associées à une moindre chance de survie (Leenhouwers et al., 2002). En effet, un tractus gastro-intestinal plus lourd peut refléter un tractus plus développé et donc plus favorable à la digestion des nutriments en période postnatale. Quant au glycogène musculaire, c'est une source d'énergie qui permet au porcelet de lutter contre l'hypothermie postnatale avant la prise colostrale (Le Dividich et al., 1998). Cependant, de nombreux autres marqueurs de la maturité néonatale mesurés dans le plasma, le muscle et le foie des nouveau-nés, ainsi que les indicateurs du stress oxydatif, n'ont pas été affectés par le milieu d'élevage des truies pendant la gestation.
Un et quatre jours après la naissance, des différences de microbiote fécal ont été observées entre les porcelets des deux milieux mais elles semblaient trop modestes pour expliquer les différences de mortalité (Kubasova et al., 2017). Quatre jours après la naissance, des différences du statut redox sont apparues chez les porcelets. Elles indiquaient que la capacité antioxydante systémique avait moins augmenté chez les porcelets nés des truies du milieu conventionnel que chez ceux issus du milieu enrichi. Dans le lait prélevé à quatre jours de lactation, le nombre de cellules a été trois fois plus faible pour les truies issues de l'environnement conventionnel, sans que les proportions des différentes populations de cellules immunitaires diffèrent (Merlot et al., 2019). Ces mesures ont été réalisées après la période critique des 72 h post-partum pendant laquelle les différences de mortalité s'exprimaient. Elles indiquent néanmoins que les conditions d'élevage des truies pendant la gestation peuvent influencer les transitions physiologiques en œuvre au niveau de la glande mammaire à la mise bas et à l'établissement de la lactation, ou au niveau des porcelets lors de leur adaptation à la vie extra-utérine.
En conclusion, en comparaison avec le milieu conventionnel, enrichir le milieu des truies gestantes en leur fournissant une litière de paille et plus d’espace a réduit leur niveau de stress pendant la gestation et a légèrement amélioré leur état de santé. Les différences de composition du colostrum et de comportement des truies après la mise bas étaient modestes. Néanmoins, cet enrichissement était aussi associé à des indicateurs de plus grande maturité des porcelets à la naissance. Il est possible que la meilleure survie des porcelets observée avec l’enrichissement du milieu pendant la gestation ait résulté de la conjonction de ces effets positifs, dont chacun pris séparément était probablement trop faible pour induire à lui seul les différences observées.
1.2. Une stratégie d’enrichissement du milieu d’élevage pour réduire le stress de la truie
Dans une seconde expérience, nous avons cherché à vérifier si les différences de taux de mortalité des nouveau-nés étaient dues à la diminution du stress des mères par l’enrichissement du milieu pendant la gestation. Pour cela, nous avons enrichi le milieu conventionnel de l’étude précédente par des objets manipulables (des cubes de bois fixés à des chaînes) et en distribuant des granulés de paille dans l’auge tout au long de la gestation. Le choix de fournir des « jouets » et des granulés de paille reposait sur l’hypothèse que la paille présente dans le milieu enrichi avait permise aux truies d’exprimer leur comportement d’investigation et de réduire leur frustration alimentaire causée par le rationnement pendant la gestation. Nous avons aussi fait l’hypothèse que ces enrichissements seuls, sans réduction de la densité des truies, permettraient de réduire le stress des truies. Les trois environnements, Conventionnel (C), Conventionnel Enrichi (CE) et Enrichi (E), présentaient ainsi des niveaux d’enrichissement croissants. Comme dans l’expérience précédente, toutes les truies étaient transférées en maternité à 105 jours de gestation, dans des stalles individuelles similaires, puis conduites de la même façon.
L'enrichissement du milieu avec les « jouets » et les granulés de paille n'a pas eu d'effet négatif sur l'état corporel et le statut métabolique des truies pendant la gestation par rapport au milieu conventionnel (Quesnel et al., 2019b). Il était important de vérifier ce point car les composés fibreux, dont la paille est riche, peuvent influencer l'absorption des nutriments et réduire la quantité d'énergie nette apportée par la ration. En revanche, les truies de l'environnement CE avaient des concentrations salivaires en cortisol intermédiaires à celles des truies des deux autres milieux (figure 3D). De plus, le comportement d'investigation des truies vis-à-vis des substrats disponibles (ATOL_0000845) augmentait avec le degré d'enrichissement du milieu alors que les activités stéréotypées (ATOL_0000852) diminuaient (figure 4, Quesnel et al., 2018). Enfin, la mortalité très précoce des porcelets (à la naissance et dans les 12 h post-partum) était plus faible dans les deux milieux enrichis (CE et E) que dans le milieu conventionnel (6,6 et 6,3 contre 11,1 % des porcelets nés totaux, figure 3C). L'enrichissement du milieu par des objets manipulables et des granulés de paille pendant la gestation a donc bien réduit le niveau de stress des truies ainsi que la mortalité très précoce des porcelets.
Figure 4. Activité comportementale des truies à 101 jours de gestation en fonction de l’enrichissement du milieu d’élevage pendant la gestation.

Milieu conventionnel (C), conventionnel enrichi avec des jouets en bois et des granulés de paille (CE) et milieu enrichi (E). L’activité a été mesurée pendant 4 h (8 h 30-10 h 30 et 13 h 30-15 h 30, observation par scan de 7 min). Moyenne ± erreur standard ; a,b,c : différences significatives au seuil de signification P < 0,05.
En conclusion, ce travail fait émerger des pistes intéressantes pour les acteurs de la filière porcine. Améliorer le bien-être des truies pendant la gestation est bénéfique aux truies mais permettrait également de réduire la mortalité précoce des porcelets. Réaménager les élevages selon le milieu le plus favorable (sur litière profonde et à densité animale réduite) n’étant pas toujours envisageable à court terme, des stratégies alternatives doivent être développées en attendant. La stratégie correctrice proposée ici, à savoir l’enrichissement du milieu conventionnel, mériterait d’être étudiée à plus grande échelle.
2. Nouvelles pistes et stratégies alimentaires pour atténuer les effets du sevrage chez le porcelet
2.1. Le sevrage : une période à haut risque pour la santé des porcelets
En production porcine conventionnelle, le sevrage est réalisé à trois ou quatre semaines d'âge. Il correspond à la séparation de la mère, au mélange avec des animaux d'autres portées, au passage du lait à un aliment en général sec et à base de céréales ainsi qu'à un changement d'environnements physique et microbien (de la case de maternité à la case de post-sevrage). Le sevrage est particulièrement stressant pour les porcelets dont les systèmes digestif et immunitaire sont encore immatures. Ces stress se traduisent souvent par une anorexie voire une période de jeûne, considérée comme l'une des premières causes de désordres digestifs d'origine alimentaire ou infectieuse (Lallès et al., 2007 ; Lallès et Guillou, 2014 ; Molist et al., 2014). Ces problèmes digestifs multifactoriels sont regroupés sous le terme de syndrome post-sevrage. Ils se caractérisent par des diarrhées (AHOL_0003105), une mortalité (AHOL_0003083) généralement faible (1,5 à 2 % ; Fairbrother et Nadeau, 2019) et par des ralentissements de la croissance (AHOL_0003101), pas toujours compensés lors des phases d'élevage ultérieures, ce qui conduit à accroitre l'hétérogénéité de poids entre les animaux et génère des pertes économiques au niveau des élevages (tableau 1). Les éleveurs traitent parfois les porcelets affectés par des antibiotiques. La forte réduction de l'utilisation globale des antibiotiques pendant la période de post-sevrage observée depuis quelques années, avec une baisse de 70 % du nombre de jours de traitements par animal entre 2010 et 2016 (Hémonic et al., 2019), a été rendue possible par la mise en place d'un ensemble de pratiques visant à limiter le stress (confort et hygiène du logement, gestion de l'allotement), renforcer la robustesse des animaux (vaccination…), et sécuriser les conduites alimentaires (optimisation du niveau d'ingestion, qualité des matières premières, valeur nutritive des aliments, additifs visant à sécuriser la santé) (Rhouma et al., 2017 ; Liu et al., 2018). Cette approche préventive et intégrée de la santé vise à renforcer la résistance des animaux, en diminuant les effets négatifs du sevrage, et à favoriser leur résilience, c'est-à-dire leur capacité à récupérer après ce stress. Dans le cadre du projet PROHEALTH, nous avons contribué à mieux comprendre les effets de deux leviers, la génétique et l'alimentation, sur la capacité des porcelets à répondre aux stress du sevrage.
2.2. Mise en évidence d’un effet génétique sur la dynamique de la réponse au sevrage
Dans un premier essai, nous avons étudié les réponses au sevrage de porcs issus des deux lignées divergentes sur la CMJR (Montagne et al., 2021), issues du dispositif de sélection présenté dans l'introduction. Notre hypothèse était que la sélection sur l'efficacité alimentaire a pu affecter la capacité d'adaptation des animaux au sevrage. Les animaux des deux lignées (n = 32 par lignée) ont été sevrés à 28 jours d'âge, logés dans des cases individuelles et ont reçu successivement deux aliments commerciaux : un aliment de démarrage (du sevrage à 11 jours après le sevrage) puis un aliment de post-sevrage (de 12 jours à trois semaines après le sevrage). Les consommations moyennes journalières d'aliment (ATOL_0005508) ainsi que les vitesses de croissance mesurées par le Gain Moyen Quotidien ou GMQ (ATOL_0000989) des porcs de la lignée CMJR-, la plus efficace, étaient respectivement 35 et 40 % plus faibles que celles des animaux de la lignée CMJR+, moins efficace, lors de la première semaine suivant le sevrage. Cette différence est à mettre en relation avec une digestibilité totale apparente de la matière organique (ATOL_0001234) inférieure de 6 %. Par ailleurs, un déficit nutritionnel plus marqué est observé chez les animaux de la lignée CMJR- pendant cette période. Chez ces derniers, les concentrations plasmatiques en acides gras libres et urée, respectivement marqueurs de mobilisation des réserves lipidiques et protéiques en réponse à une sous-nutrition, étaient 167 et 55 % plus élevées par rapport aux animaux de la lignée CMJR+. Lors de la première semaine suivant le sevrage, la prévalence des diarrhées était plus importante pour la lignée CMJR- (12 porcelets pour les CMJR- vs un pour les CMJR+), ces animaux présentant aussi une concentration en haptoglobine plasmatique supérieure (+ 52 %) comparativement aux CMJR+. Chez ces mêmes porcelets CMJR-, l'abondance de certaines espèces des genres Fusobacterium ou Escherichia dans les fèces, caractéristiques du microbiote des porcelets allaités, a diminué plus lentement après le sevrage comparativement aux porcelets CMJR+. L'évolution plus lente des populations microbiennes fécales après le sevrage peut expliquer la différence de prévalence des diarrhées (Kubasova et al., 2018). Certaines souches pathogènes d'Escherichia coli (AHOL_0004068) présentes dans l'intestin auraient ainsi bénéficié de plus de temps pour se développer et favoriser l'apparition de diarrhées infectieuses (Fairbrother et al., 2005). Au cours des deuxième et troisième semaines après le sevrage, les animaux des deux lignées avaient les mêmes performances zootechniques et des caractéristiques physiologiques similaires. Cette absence de différence se maintenait jusqu'à la fin de la période de post-sevrage. Cette étude a mis ainsi en évidence des différences dans la dynamique de réponse au sevrage selon la lignée génétique. Dans nos conditions expérimentales, les animaux de la lignée avec une efficacité alimentaire élevée (CMJR-) se sont avérés être moins résistants au stress lors de la semaine qui suit le sevrage mais être aussi résilients car capables de récupérer rapidement après le sevrage, rattrapant les porcelets de la lignée CMJR+.
2.3. Mise en évidence d’une interaction entre stratégie alimentaire et lignée génétique
Comme la génétique, l'alimentation est un facteur de modulation des réponses des porcelets au sevrage. Dans un second essai, 132 porcs de chacune des lignées CMJR ont été logés collectivement de 28 jours à 10 semaines d'âge. Ils ont reçu en post-sevrage soit la même séquence alimentaire que dans l'essai précédent (séquence témoin avec deux aliments successifs), soit une séquence alimentaire plus complexe comprenant trois aliments successifs et considérée comme sécurisante par la composition en matières premières (qualité des céréales et des protéines), leurs valeurs nutritives, et la présence de pulpe de chicorée et d'extraits de plantes (flavonoides) (Gilbert et al., 2019). Tous les animaux ont ensuite reçu le même aliment commercial en période de croissance-finition entre 10 et 23 semaines d'âge. Nous avons confirmé les différences entre les deux lignées observées dans le premier essai, avec une consommation d'aliment et une vitesse de croissance mesurées immédiatement après le sevrage plus faibles (figure 5), et un déficit nutritionnel plus marqué chez les animaux de la lignée CMJR-. La proportion d'individus présentant des diarrhées a été aussi plus élevée chez ces animaux de la lignée CMJR- (22 contre 14 % pour la lignée CMJR+) mais les concentrations en haptoglobine plasmatique ne différaient pas. Les porcelets ayant reçu les aliments de la séquence complexe ont consommé plus d'aliment et ont eu une meilleure croissance (+ 5 % par rapport à l'aliment témoin en moyenne pour les deux lignées confondues) et moins de diarrhées (12 vs 24 % des animaux). Ces effets ont été plus marqués chez les animaux de la lignée CMJR- (figure 5).
Figure 5. Dynamique d’adaptation au sevrage des lignées CMJR+ et CMJR- en fonction de la séquence alimentaire en post-sevrage.

Influence de la lignée (CMJR+ ou CMJR-) et de la séquence alimentaire (témoin ou sécurisante) sur la dynamique de la réponse des porcelets après le sevrage à 28 jours d’âge. Les semaines 1 et 3 correspondent aux semaines après le sevrage.
L’utilisation de la séquence alimentaire complexe a ainsi pu limiter les conséquences du sevrage chez les animaux CMJR- (figure 5) mais n’a pas eu d’effet à long terme, lors de la période de croissance et finition entre 10 et 23 semaines d’âge. En pratique, ces résultats confirment l’intérêt de sécuriser l’alimentation des porcelets lors de cette période critique du sevrage et plus particulièrement celle des animaux les plus sensibles.
3. Vers une meilleure compréhension de l’effet des conditions de logement sur la santé du porc en croissance en interaction avec la lignée génétique
3.1. Une forte prévalence des lésions d’ostéochondrose chez le porc en croissance
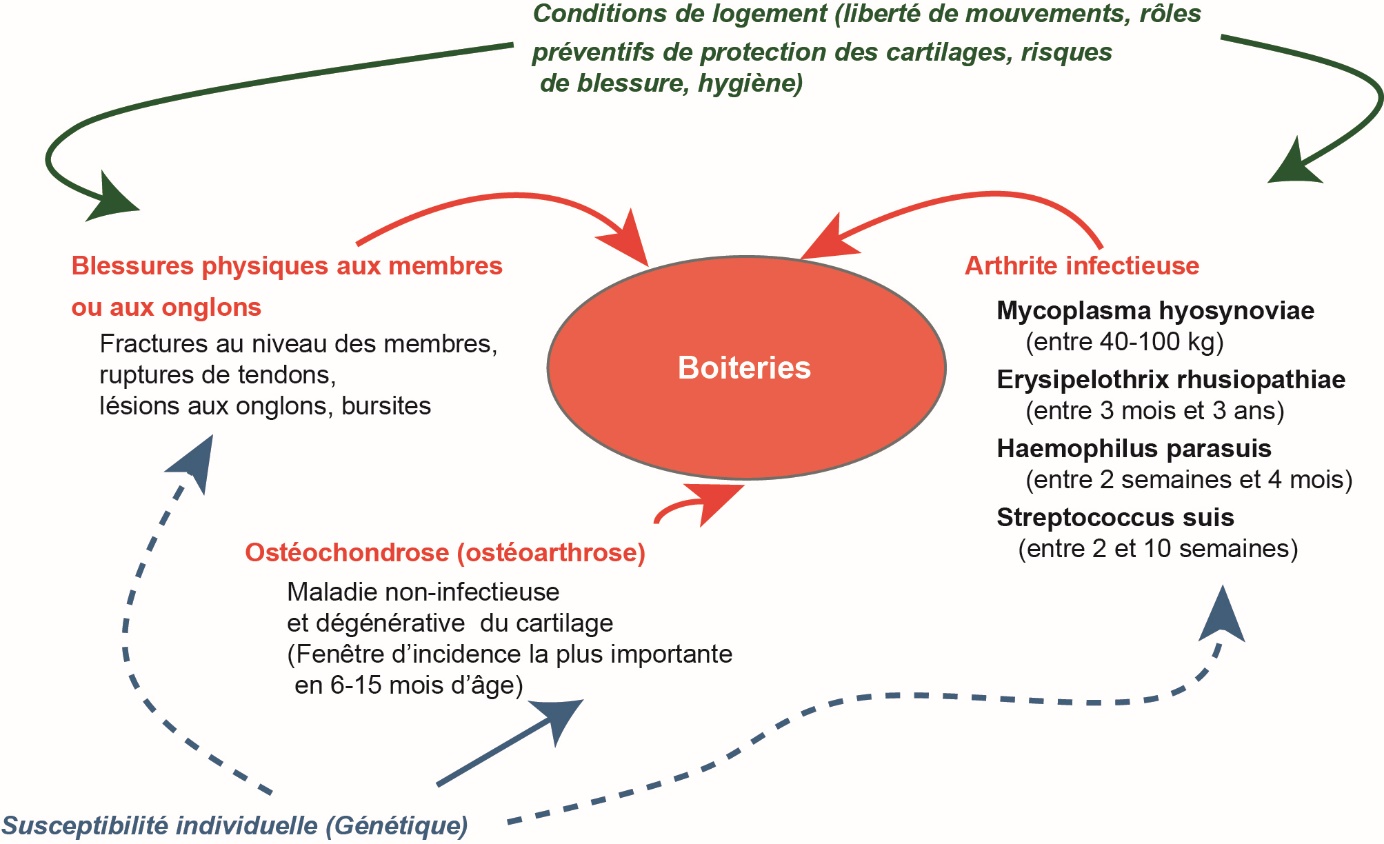
Les troubles locomoteurs sont le plus souvent détectés par la présence de boiteries (AHOL_0003029) ou par l'examen des articulations à l'abattoir (Etterlin et al., 2014 ; Stavrakakis et al., 2014). Les boiteries sont le symptôme le plus visible de ces troubles. Elles sont responsables de 10 à 20 % des réformes chez les truies, ce qui en fait la seconde ou la troisième cause de réforme (Stavrakakis, 2014). Dans les populations de porcs en croissance, la prévalence des boiteries se situe entre 2 et 21 % (Stavrakakis, 2014) mais se traduit peu en réforme du fait de la faible durée de vie de ces animaux. Les troubles locomoteurs représentent une atteinte au bien-être animal quel que soit l'âge des animaux, car la douleur est souvent la cause première des boiteries. Ils sont aussi une cause importante de pertes économiques, du fait des frais vétérinaires (tableau 1), mais aussi des réformes prématurées qu'ils induisent. Un facteur de risque des boiteries (Jensen et Toft, 2009 ; figure 6) est l'ostéochondrose (OC ; AHOL_0005230), une maladie non infectieuse et dégénérative correspondant à une nécrose ischémique du cartilage de croissance et dont l'incidence est particulièrement forte entre six et 15 mois (van Grevenhof et al., 2011 ; Etterlin et al., 2014). Pour les animaux reproducteurs, la survenue d'ostéochondrose à un jeune âge peut être une cause majeure de boiteries à l'âge adulte. La prévalence de cette maladie est très élevée dans les races de porcs les plus courantes (Stavrakakis et al., 2014). Il est établi que la race ou la lignée génétique sont les facteurs de prédisposition les plus importants (Petersen et al, 1998 ; Etterlin et al., 2015). Par ailleurs, un débat existe quant à un lien entre la vitesse de croissance et la prévalence de l'OC qui n'est pas observé de façon homogène selon les études (Lundeheim, 1987 ; Ytrehus et al., 2007). Nous avons donc évalué la prévalence de cette maladie dans les expérimentations conduites sur les porcs en croissance du projet PROHEALTH, sur les deux lignées sélectionnées sur la CMJR. Nos résultats montrent une très forte prévalence de l'OC sur tous les animaux lors de l'abattage à 100 kg de poids vif, soit à un âge de 21 à 22 semaines, avec 32 à 82 % des porcs atteints selon les articulations étudiées (cartilages des extrémités distales et proximales des fémurs et humérus). Même si les résultats varient considérablement entre les études, ces chiffres confirment la forte prévalence de ce type de lésions dans les races de porcs domestiques (Lundeheim, 1987 ; Jørgensen et Andersen, 2000 ; van Grevenhof et al., 2011 ; Etterlin et al., 2014). Notre étude a également montré que les porcs de la lignée CMJR- étaient plus nombreux à présenter des lésions d'OC, en particulier au niveau des épaules (extrémité proximale de l'humérus) et des hanches (extrémité proximale du fémur). Ces résultats sont cohérents avec leur poids et la teneur en viande maigre des carcasses plus élevés et leur vitesse de croissance plus rapide dans cette étude. En effet, certaines observations de la littérature concluent à un lien entre la vitesse de croissance, la teneur en viande maigre des carcasses, et la prévalence des lésions d'OC (Lundeheim, 1987 ; Jørgensen et Andersen, 2000). Nos résultats confirment le rôle de la génétique dans la susceptibilité à ce type de maladie. Malgré cela, nous avons détecté peu de porcs réellement boiteux. La démarche des porcs CMJR- a seulement été jugée comme marginalement dégradée, avec des scores laissant entrevoir une démarche plus raide. Ceci illustre qu'une partie importante des lésions d'OC est peu symptomatique jusqu'à un poids vif de 100 kg.
Figure 6. Facteurs de risque des boiteries chez le porc : multiples et dépendants de l'âge de l’animal (d’après Jensen et Toft, 2009).
Nous avons testé la possibilité de détecter précocement l'OC sans recours à l'abattage ou à l'utilisation d'appareil d'imagerie onéreux, par analyse de biomarqueurs sériques de la synthèse ou de la dégradation du collagène de type II constitutif des cartilages (respectivement CPII, propeptide du procollagène de type II et C2C protéine de clivage du collagène de type II ; Frantz et al., 2010). Les teneurs sériques en ces biomarqueurs ont bien été plus élevées dans la lignée CMJR- présentant une plus grande susceptibilité à l'OC, ce qui est cohérent avec le fait que l'OC peut se traduire par une plus grande dégradation des cartilages mais aussi plus de synthèse du fait des processus de réparation qui peuvent être induits (Frantz et al., 2010). Les teneurs sériques en ces biomarqueurs ne peuvent néanmoins pas être utilisées pour une détection individuelle de la maladie car nous n'avons observé aucune corrélation entre ces teneurs et les scores individuels d'ostéochondrose.
3.2. L’activité physique : un levier pour limiter les troubles locomoteurs à ajuster selon la lignée génétique
Une restriction de la liberté de mouvement et la présence de sols durs et abrasifs sont des conditions courantes dans les bâtiments d'engraissement. Les préoccupations de la société concernant le bien-être animal incitent à favoriser l'augmentation des surfaces disponibles pour les animaux et l'enrichissement des milieux d'élevage porcin, ce qui augmente les possibilités de mobilité et d'activité physique (EFSA, 2014). Il a été montré chez le porc en croissance qu'une activité physique modérée a un effet bénéfique sur la prévention des troubles locomoteurs (Perrin et Bowland, 1977 ; Petersen et al., 1998), mais aussi qu'une activité physique trop importante peut avoir des effets délétères (Schenck et al., 2008). Dans le projet PROHEALTH, nous avons cherché à déterminer si les bénéfices d'une activité physique accrue, dans le cas de porcs en croissance élevés sur sol dur, différaient pour les deux lignées sélectionnées sur la CMJR. Meunier-Salaün et al. (2014) avaient rapporté dans une étude précédente une plus faible sensibilité aux troubles locomoteurs des porcs de la lignée CMJR- relativement aux porcs CMJR+ sur des effectifs limités de porcs.
Dans notre étude, 160 porcs, dont 80 CMJR- et 80 CMJR+, étaient logés en un groupe sur un sol bétonné sans litière et nourris grâce à des distributeurs automatiques d'aliment (DACs) auxquels ils accédaient par un système de trieur. Le trieur, grâce à l'identification individuelle des porcs (puces RFID), permettait d'augmenter l'activité physique de la moitié des porcs de chaque lignée, en leur faisant parcourir deux fois toute la longueur de la salle avant d'accéder à nouveau aux DACs en sortie de trieur. Ce traitement (A+) a été comparé à un traitement témoin qui concernait l'autre moitié des porcs, ces derniers pouvant accéder à nouveau aux DACs dès leur sortie du trieur (témoin). Le traitement A+ a bien conduit à une activité physique effective accrue (mesurée par le nombre quotidien de passages dans le trieur), surtout chez les porcs CMJR+ (figure 7A) pour lesquels on avait déjà caractérisé une activité physique en moyenne plus importante (Gilbert et al., 2017). Tous les animaux n'auraient donc pas les mêmes capacités ou la même motivation à augmenter leur activité physique et la distance parcourue pour obtenir de la nourriture. Malgré une prévalence des boiteries faibles dans cet essai, l'augmentation de l'activité physique a permis une légère diminution du score moyen de démarche chez les porcs du traitement A+ par rapport aux porcs du lot témoin (scores de 0,42 contre 0,55, P < 0,05, la note de 0 correspondant à une démarche souple et celle de 5 à des animaux incapables de se déplacer). Cette diminution était surtout présente pour les porcs CMJR+ (0,51 contre 0,33, figure 7B) sans que l'interaction traitement × lignée n'ait été significative (P < 0,10).
Figure 7. Dynamique d’adaptation des lignées CMJR+ et CMJR- à une augmentation induite de leur activité physique (A) et score de démarche moyen sur l’essai (B).

Influence de la lignée (CMJR- ou CMJR+) et du traitement expérimental (trait, T = témoin et A+= activité physique induite) sur la dynamique de l’activité physique effective des porcs en croissance mesurée par leur nombre de passages journaliers au trieur et sur les scores de démarche (n=160). *** : P < 0,001 ; ** : P < 0,01 ; * : P < 0,05 ; t : P < 0,10 ; ligne haute : effet de la lignée ; ligne médiane : effet du traitement, ligne basse : effet de l’interaction. Scores de démarche entre 0 et 5 : 0 démarche fluide, 5 déplacement impossible.
3.3. Les conditions d’hygiène du logement affectent durablement la santé et la croissance des porcs
En élevage porcin, le respect des bonnes pratiques d'hygiène est primordial pour prévenir les risques sanitaires liés à la présence d'agents abiotiques et biotiques (Corrégé et al., 2011). En France et en Europe, les bonnes pratiques d'hygiène sont globalement moins bien respectées dans les élevages de production que dans les élevages de sélection (Gueriaux et al., 2017). Au sein d'un même élevage, les protocoles de nettoyage et de désinfection sont moins souvent appliqués en engraissement qu'en post-sevrage et en maternité (Hémonic, 2010). Une marge de progrès est donc encore possible notamment pour réduire la prévalence des maladies respiratoires.
En effet, en période d'engraissement, et particulièrement en élevage en claustration, les maladies respiratoires sont depuis longtemps très fréquentes (Madec et Kobisch, 1982 ; Fablet et al., 2013). Des études plus récentes indiquent que cela reste le cas malgré le déploiement de la vaccination contre le mycoplasme (Krejci et al., 2017 ; Krejci et al., 2018). Les maladies respiratoires ont une origine multifactorielle et impliquent des agents infectieux et des facteurs non infectieux comme la densité en poussières et la concentration en gaz et en endotoxines bactériennes dans les salles d'élevage. D'ailleurs, la prévalence des atteintes pulmonaires est plus élevée dans les élevages où les règles d'hygiène et de biosécurité ne sont pas correctement respectées (Fablet et al., 2013).
Dans le cadre du projet européen PROHEALTH, nous avons évalué les conséquences du non-respect de l’hygiène du logement de 12 à 18 semaines d’âge (début de la période d’engraissement) sur la santé et les performances de croissance des porcs jusqu’à 26 semaines d’âge. Pendant cette période, 160 porcs issus des lignées CMJR+ et CMJR- (n = 80 pour chaque lignée) ont été élevés pour moitié en conditions d’hygiène dégradée, et pour moitié en bonnes conditions d’hygiène. Les conditions d’hygiène dégradées consistaient en l’absence de nettoyage de la salle d’élevage et de vide sanitaire entre bandes, et au mélange avec des porcs non expérimentaux issus de bandes d’élevage différentes. La ventilation était également réduite. Les valeurs médianes de teneurs en dioxyde de carbone (1 186 vs 1 060 ppm) et ammoniac (3,5 vs 7,25 ppm) dans l’air ainsi que les températures ambiantes (21 vs 20,7 °C), étaient globalement similaires dans les conditions optimales et dégradées. Ces valeurs étaient inférieures aux seuils de 1 540 ppm pour le dioxyde de carbone et de 11 ppm pour l’ammoniac considérées comme critiques pour la santé des porcs (Murphy, 2011).
Notre étude a montré que, pour les deux lignées de porcs, le non-respect des règles d'hygiène dégradait les performances de croissance. Le ralentissement de la vitesse de croissance (GMQ) de 20 % (figure 8) avait peu d'incidence sur la composition corporelle, mais était associé à une augmentation du nombre de porcs présentant des lésions pulmonaires de type pneumonie (AHOL_0003040) et de pleurésie. Malgré l'absence de différence sur les teneurs en gaz, l'humidité relative, plus élevée dans l'environnement aux mauvaises conditions d'hygiène (74,5 vs 54,5 %), pourrait avoir favorisé la pression bactérienne de sorte que les porcs auraient été exposés à une plus large gamme de contaminants microbiens et d'endotoxines présents dans l'atmosphère (Banhazi et al., 2008). La moitié des porcs ont été abattus à l'issue de ces six semaines en conditions d'hygiène contrastées, les porcs restants étant transférés dans de bonnes conditions d'hygiène jusqu'à la fin de l'essai. La poursuite de l'engraissement en bonnes conditions d'hygiène jusqu'à l'abattage à l'âge de 26 semaines n'a pas permis aux porcs précédemment élevés en conditions dégradées de compenser ce ralentissement de la croissance, puisque leur poids vif à un même âge d'abattage restait inférieur de près de 10 kg en moyenne à celui des porcs élevés précédemment en bonnes conditions d'hygiène (figure 8A et 8B) ; leur indice de consommation a cependant été amélioré pendant cette seconde période bien que l'indice global sur les deux périodes restait dégradé. La qualité de la viande (pH : ATOL_0001684, couleur : ATOL_0001017, perte en eau de la longe) en fin d'engraissement n'a pas été significativement affectée par les conditions d'hygiène en début d'engraissement.
Figure 8. Influence des conditions d’hygiène du logement sur la croissance et des indicateurs sanguins d’inflammation chez le porc en croissance.
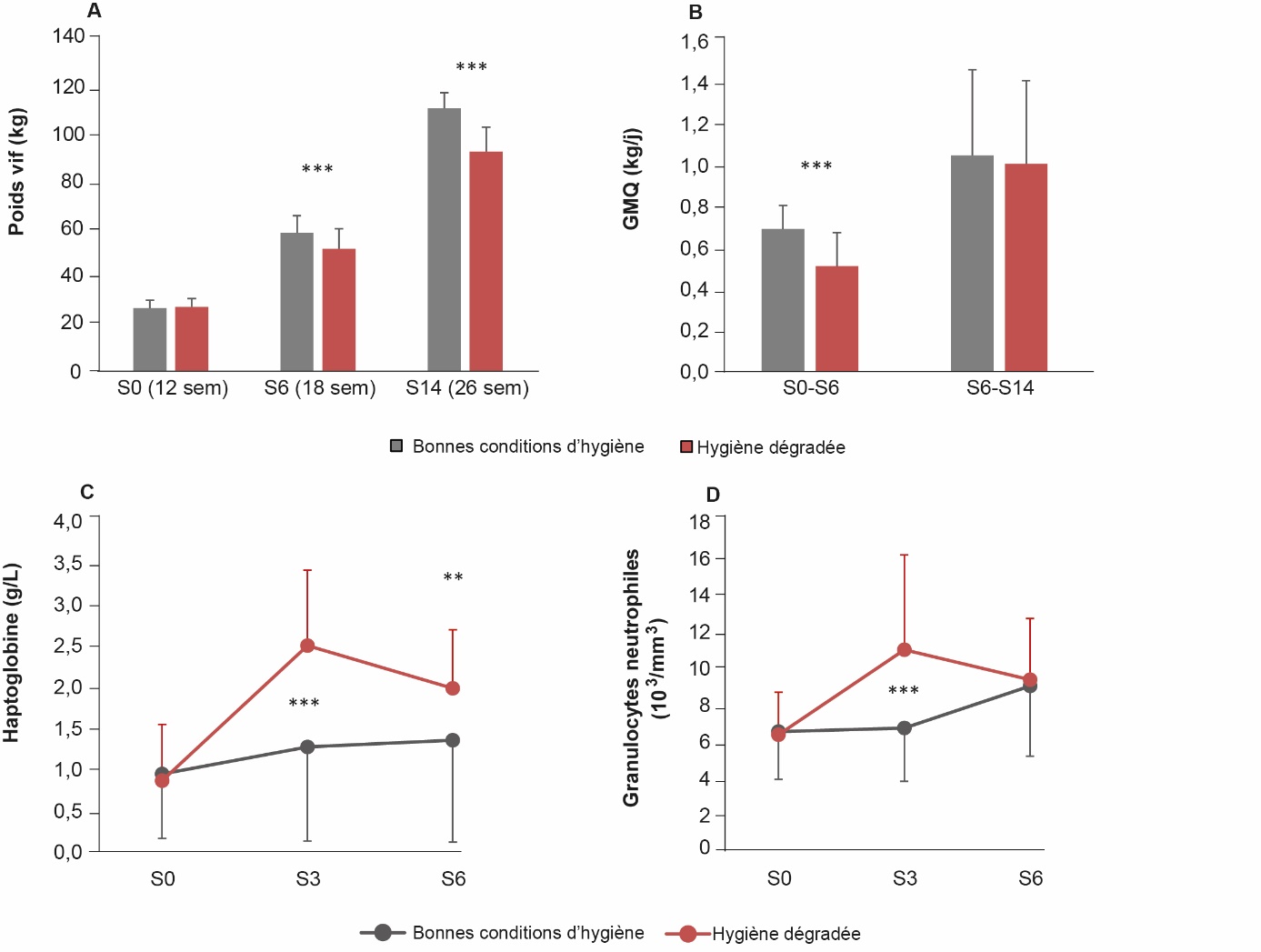
Les porcs (n=160) ont été élevés pendant six semaines (de S0 à S6, soit entre l’âge de 12 et 18 semaines) en conditions d’hygiène dégradée ou dans de bonnes conditions d’hygiène, puis en bonnes conditions d’hygiène pendant huit semaines supplémentaires (S6-S14, soit entre l’âge de 18 et 26 semaines). Les poids vifs (A) ont été mesurés à jeun au début de l’essai (S0), six semaines (S6) et 14 semaines (S14) plus tard. Le gain de poids moyen quotidien (GMQ, B) a été calculé entre S0 et S6 puis de S6 à S14. Des prises de sang ont été effectuées à jeun. L’haptoglobine (C), une protéine inflammatoire, a été dosée dans le plasma alors que la quantité de granulocytes (D) a été mesurée dans le sang. Pour ces deux variables, seules les données mesurées lors de la période 1 sont présentées. *** P < 0.001 et ** P < 0.05.
Nous avons également étudié les mécanismes physiologiques à l'origine des problèmes de santé et des moindres performances en réalisant sur l'ensemble des porcs des prélèvements de sang, de salive et de fèces en élevage, et de tissus à l'abattoir. Les concentrations plasmatiques en haptoglobine ont été doublées et le nombre de granulocytes a augmenté de 60 % après trois semaines en conditions dégradées (figure 8C et 8D). Les teneurs en immunoglobulines sanguines, celles d'indicateurs du stress oxydant tissulaire (équilibre entre capacité anti-oxydante et production d'hydro-peroxydes) et les activités des enzymes anti-oxydantes dans les tissus adipeux étaient également plus élevées en conditions dégradées (Sierźant et al., 2019). Au-delà des réponses moyennes contrastées, cette étude a également montré une variabilité interindividuelle des concentrations sanguines en 51 nutriments et indicateurs immunitaires, et des performances des animaux. Cette variabilité a été mise à profit pour identifier, à l'aide de régressions linéaires multiples six variables permettant d'expliquer la vitesse de croissance journalière. Elles correspondent aux variations des concentrations plasmatiques au cours des trois premières semaines d'élevage en conditions d'hygiène dégradée (Le Floc'h et al., 2021) de variables indicatrices du statut inflammatoire (haptoglobine), de la capacité anti-oxydante du plasma, et du métabolisme (acides gras libres, trois acides aminés : τ-méthylhistidine, isoleucine et tryptophane). Ces résultats soulignent, d'une part, le rôle clé du métabolisme dans l'expression des performances et, d'autre part, les interrelations entre métabolisme et statut inflammatoire. À terme, cette approche devra être validée sur d'autres jeux de données et enrichie par la mesure d'autres indicateurs pour proposer des biomarqueurs prédictifs de la vitesse de croissance. Enfin, notre étude a permis de suggérer le rôle de certains métabolites plasmatiques, comme la τ-méthylhistidine, dont le rôle est encore peu décrit dans la littérature sur le porc.
Ce travail a également montré que les deux lignées divergentes pour la CMJR ne se comportaient pas de la même façon face à ces conditions d'élevage dégradées (Chatelet et al., 2018) : la croissance de la lignée CMJR- était moins affectée à la fin des six semaines d'élevage en hygiène dégradée (669 vs 478 g/j pour les CMJR- et CMJR+, respectivement) avec, à l'âge d'abattage (25 semaines), une différence de poids vif de 5,5 et 13,4 kg entre conditions d'hygiène pour respectivement les CMJR- et CMJR+. La réponse inflammatoire (haptoglobine) et le stress oxydant étaient également plus faibles dans la lignée CMJR- soumise aux conditions d'hygiène dégradée (Sierźant et al., 2019). Enfin, la prévalence et la sévérité des lésions pulmonaires étaient plus faibles pour la lignée CMJR-. Contrairement aux études portant sur le sevrage (cf 2.1) et l'ostéochondrose (cf 3.1), la lignée ayant une efficacité alimentaire plus élevée (CMJR-) s'est montrée plus résistante aux conditions d'hygiène dégradée que celle dont l'efficacité est plus faible. Une étude complémentaire a montré des différences dans le métabolisme postprandial de certains acides aminés comme la lysine et le tryptophane entre les deux lignées. De telles différences pourraient expliquer la meilleure capacité des porcs de la lignée CMJR- à faire face à une mauvaise hygiène de leurs conditions de logement (Fraga et al., 2021). En effet, le tryptophane, un acide aminé dont le métabolisme est impliqué dans la régulation du système immunitaire (Le Floc'h et al., 2011) voit ses concentrations plasmatiques non modifiées par la dégradation de l'hygiène chez les CMJR+, alors que l'accumulation de la lysine dans le plasma résulte de l'effet plus important des conditions d'hygiène dégradée sur la croissance des porcs CMJR+, cet acide aminé étant moins incorporé dans les protéines musculaires.
En conclusion, ce travail a souligné l’importance du respect des règles d’hygiène dans les bâtiments d’élevage pour préserver la croissance et la santé des porcs tout au long de la phase d’engraissement. Du point de vue économique, le non-respect des conditions d’hygiène entrainant un ralentissement du gain de poids de près de 20 % pourrait réduire la marge par porc de 15 à 25 euros par rapport à celle obtenue avec des porcs élevés dans de meilleures conditions d'hygiène (Le Floc’h et Niemi, 2018). Enfin, la lignée génétique influence également la réponse des porcs à ces conditions d’élevage délétères pour la santé et les performances.
Conclusion
Ce vaste projet a mis en exergue l’importance des conditions et pratiques d’élevage, incluant l’alimentation, dans la sensibilité des porcs aux maladies de production et souligne les efforts à poursuivre pour développer des élevages plus vertueux dans la prise en compte de la santé et du bien-être des animaux. Une approche intégrée est nécessaire pour prévenir ces maladies multifactorielles par l’adaptation de la conduite des élevages. Grâce au modèle expérimental des lignées divergentes sur la CMJR, nous avons également montré que la susceptibilité des porcs à différents stress ou maladies est influencée par la génétique bien que la différence de susceptibilité entre lignées varie selon les maladies et les variables de réponse étudiées. La sélection génétique sur des critères de santé ou le choix de races ou de lignées plus adaptées aux conditions ou systèmes d’élevage sont aussi des pistes à développer. Au niveau individuel, disposer d’indicateurs précoces (éventuellement intégratifs) en s’appuyant notamment sur la diversité des réponses biologiques mobilisées par les animaux, permettrait d’identifier rapidement les animaux candidats à des traitements ciblés.
Remerciements
Nous remercions nos collègues des UMR PEGASE (Physiologie, Environnement et Génétique pour l'Animal et les Systèmes d'Élevage) et UMR ISP (Infectiologie et Santé Publique), des unités expérimentales UE3P (Physiologie et Phénotypage des Porcs ; https://doi.org/10.15454/1.5573932732039927E12) et UE GENESI (Génétique, Expérimentation et Système Innovants ; https://doi.org/10.15454/1.5572415481185847E12 ), du groupe CCPA et de la Chambre Régionale d’Agriculture de Bretagne (CRAB) ayant participé au travail et nos partenaires européens (Veterinary Research Institute (VRI) en République Tchèque et l’institut finlandais Luke).
Notes
- Nathalie Le Floc’h, Anne Boudon (anne.boudon@inrae.fr), Lucile Montagne (lucile.montagne@agrocampus-ouest.fr) et Hélène Quesnel (helene.quesnel@inrae.fr) sont les principales rédactrices.
- Les ontologies ATOL et AHOL sont respectivement deux ontologies des caractères phénotypiques de production (Animal Trait Ontology for Livestock) et des caractères de santé des animaux d’élevage (Animal Health Ontology for Livestock). Dans le texte, chaque caractère est référencé par son identifiant unique ATOL ou AHOL.
Références
- Banhazi T.M., Seedorf J., Rutley D.L., Pitchford W.S., 2008. Identification of risk factors for sub-optimal housing conditions in Australian piggeries: Part 2. Airborne pollutants. J. Agric. Saf. Health, 14, 21-39. https://doi.org/10.13031/2013.24122
- Baxter E.M., Rutherford K.M.D., D'Eath R.B, Arnott G., Turner S.P., Sandøe P., Moustsen V.A., Thorup F., Edwards S.A., Lawrence A.B., 2013. The welfare implications of large litter size in the domestic pig II: management factors. Anim. Welfare, 22, 219-238. https://doi.org/10.7120/09627286.22.2.219
- Cador C., Pol F., Hamoniaux M., Dorenlor V., Eveno E., Guyomarc'h C., Rose N., 2014. Risk factors associated with leg disorders of gestating sows in different group-housing systems: A cross-sectional study in 108 farrow-to-finish farms in France. Prev. Vet. Med., 116, 102-110. https://doi.org/10.1016/j.prevetmed.2014.05.004
- Chatelet A., Gondret F., Merlot E., Gilbert H., Friggens N.C., Le Floc'h N., 2018. Impact of hygiene of housing conditions on performance and health of two pig genetic lines divergent for residual feed intake. Animal, 12, 350-358. https://doi.org/10.1017/S1751731117001379
- Corrégé I., Berthelot N., Aubry A., Badouard B., Hémonic A., 2011. Biosécurité, maitrise sanitaire, conception et conduite d’élevage : impact sur les performances technico-économiques. Journ. Rech. Porcine, 43, 265-271.
- Damm B.I., Forkman B., Pedersen L.J., 2005. Lying down and rolling behaviour in sows in relation to piglet crushing. Appl. Anim. Behav. Sci., 90, 3-20. https://doi.org/10.1016/j.applanim.2004.08.008
- Eckersall P.D., Bell R., 2010. Acute phase proteins: Biomarkers of infection and inflammation in veterinary medicine. Vet. J., 185, 23-27. https://doi.org/10.1016/j.tvjl.2010.04.009
- Edwards S.A., 2002. Perinatal mortality in the pig: environmental or physiological solutions? Livest. Prod. Sci., 78, 3-12. https://doi.org/10.1016/S0301-6226(02)00180-X
- Edwards S.A., Matheson S., Pandolfi F., Thompson R., Ploetz T., Kyriazakis I., 2016. PROHEALTH, production diseases and piglet mortality. Ann Meet. Eur. Fed. Anim. Sci., Belfast, United Kingdom, 67, 314.
- EFSA, 2014. Scientific Opinion concerning a Multifactorial approach on the use of animal and non-animal-based measures to assess the welfare of pigs. EFSA Journal, 12, 3702-3802. https://doi.org/10.2903/j.efsa.2014.3702
- Etterlin P.E., Ytrehus B., Lundeheim N., Heldmer E., Osterberg J., Ekman S., 2014. Effects of free-range and confined housing on joint health in a herd of fattening pigs. BMC Vet. Res., 10, 208-221. https://doi.org/10.1186/s12917-014-0208-5
- Etterlin P.E., Morrison D.A., Österberg J., Ytrehus B., Heldmer E., Ekman S., 2015. Osteochondrosis, but not lameness, is more frequent among free-range pigs than confined herd-mates. Acta Vet. Scand., 57, 63-71. https://doi.org/10.1186/s13028-015-0154-7
- Fablet C., Dorenlor V., Eono F., Eveno E., Jolly J.P., Portier F., Bidan F., Madec F., Rose N., 2013. Facteurs non infectieux associés à la pneumonie et à la pleurésie dans 143 élevages naisseurs-engraisseurs du Grand Ouest de la France. Journ. Rech. Porcine, 45, 249-254.
- Fairbrother J.M., Nadeau E., 2019. Colibacillosis. In: Diseases of Swine, 11e Edition. Zimmerman J., Karriker L.A., Ramirez A., Schwartz K.J., Stevenson G.W., Zhang J. (Eds), John Wiley & Sons, Hoboken, New Jersey, USA, 807-834. https://doi.org/10.1002/9781119350927.ch52
- Fairbrother J.M., Nadeau E., Carlton L.G., 2005. Escherichia coli in postweaning diarrhea in pigs: an update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev., 6, 17-39. https://doi.org/10.1079/AHR2005105
- Fraga A.Z., Louveau I., Campos P.H.R.F., Hauschild L., Le Floc'h N., 2021. Selection for feed efficiency elicits different postprandial plasma metabolite profiles in response to poor hygiene of housing conditions in growing pigs. PLoS ONE, 16, e0246216. https://doi.org/10.1371/journal.pone.0246216
- Frantz N.Z., Friesen K.G., Andrews G.A., Tokach M.D., Yamka R.M., Loughin T.L., Nelssen J.L., Dritz S.S., 2010. Use of serum biomarkers to predict the development and severity of osteochondrosis lesions in the distal portion of the femur in pigs. Am. J. Vet. Res., 71, 946-952. https://doi.org/10.2460/ajvr.71.8.946
- Gilbert H., Billon Y., Brossard L., Faure J., Gatellier P., Gondret F., Labussière E., Lebret B., Lefaucheur L., Le Floch N., Louveau I., Merlot E., Meunier-Salaün M.C., Montagne L., Mormède P., Renaudeau D., Riquet J., Rogel-Gaillard C., Van Milgen J., Vincent A., Noblet J., 2017. Sélection pour la consommation alimentaire moyenne journalière résiduelle chez le porc : impacts sur les caractères et défis pour la filière. INRA Prod. Anim., 30, 439-454. https://doi.org/10.20870/productions-animales.2017.30.5.2274
- Gilbert H., Ruesche J., Muller N., Billon Y., Robert F., Montagne L., 2019. Responses to weaning in two pig lines divergently selected for residual feed intake depending on diet. J. Anim. Sci., 97, 43-54. https://doi.org/10.1093/jas/sky416
- Gueriaux D., Fediaevsky A., Ferreira B., 2017. La biosécurité : investissement d'avenir pour les élevages français. Bull. Acad. Vét. France, 170, 112-117. https://doi.org/10.4267/2042/62331
- Hémonic A., 2010. Quelles sont les pratiques de biosécurité et d'hygiène en élevages de porcs. TechniPorc. La revue technique de l'IFIP, 33, 1-13.
- Hémonic A., Poissonnet A., Chauvin C., Corrégé I., 2019. Evolution des usages d’antibiotiques dans les élevages de porcs en France entre 2010 et 2016 au travers des panels J. Rech. Porcine, 51, 278-282.
- Jensen T.B., Toft N., 2009. Causes of and predisposing risk factors for leg disorders in growing-finishing pigs. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Res., 4, 1-8. https://doi.org/10.1079/PAVSNNR20094010
- Jørgensen B., Andersen I., 2000. Genetic parameters for osteochondrosis in Danish Landrace and Yorkshire boars and correlations with leg weakness and production traits. Anim. Sci., 71, 427-434. https://doi.org/10.1017/S1357729800055442
- Knap P.W., Rauw W.M., 2009. Selection for high production in pigs. In Resource allocation theory applied to farm animal production. Rauw W.M. (Ed). CABI Editions, Wallingford, UK, 210-229. https://doi.org/10.1079/9781845933944.0210
- Kranendonk G., Mulder E.J.H., Parvizi N., Taverne M.A.M., 2008. Prenatal stress in pigs: Experimental approaches and field observations. Exp. Clin. Endocrinol. Diabetes, 116, 413-422. https://doi.org/10.1055/s-2008-1065335
- Krejci R., Gobbi S., Maynard B., Mazerolles P., 2017. Lung lesion survey using CEVA lung program in Finistère, Brittany, France. In: Proceedings of ESPHM. Nechvátalová K., Nathues H. (Eds). Europ. Symp. Porcine Health Manag. Prague, République Tchèque. 9, 307.
- Krejci R., Mazerolles P., Mortier M., 2018. Lung scoring survey in European countries in 2017. In: Proceedings of ESPHM. Segales J. (Ed). Eur. Sym. Porcine Health Manag. Barcelone, Espagne. 10, 288.
- Kubasova T., Davidova-Gerzova L., Merlot E., Medvecky M., Polansky O., Gardan-Salmon D., Quesnel H., Rychlik I., 2017. Housing systems influence gut microbiota composition of sows but not of their piglets. PLoS ONE, 12, e0170051. https://doi.org/10.1371/journal.pone.0170051
- Kubasova T., Davidova-Gerzova L., Cejkova D., Montagne L., Le Floc'h N., Rychlik I., 2018. Host genetics and environmental conditions influence microbiota composition of pigs. PLoS ONE, 13, e0201901. https://doi.org/10.1371/journal.pone.0201901
- Lallès J.P., Guillou D., 2014. Pig intestine, weaning and dietary interventions. In: Intestinal health. Key to maximise growth performance in livestock. Niewold T. (Ed). Wageningen Academic Publishers, Wageningen, Pays-Bas, 139-168. https://doi.org/10.3920/978-90-8686-792-9_6
- Lallès J.P., Bosi P., Smidt H., Stokes C.R., 2007. Nutritional management of gut health in pigs around weaning. Proc. Nutr. Soc., 66, 260-268. https://doi.org/10.1017/S0029665107005484
- Le Dividich J., Noblet J, Herpin P. van Milgen J., Quiniou N., 1998. Thermoregulation. In: Progress in Pig Science. Wiseman J., Varley M.A., Charlick J.P. (Eds). Nottingham University Press. Nottingham, UK, 229-263.
- Leenhouwers J.I., Knol E.F., de Groot P.N., Vos H., van der Lende T., 2002. Fetal development in the pig in relation to genetic merit for piglet survival. J. Anim. Sci., 80, 1759-1770. https://doi.org/10.2527/2002.8071759x
- Le Floc’h N., Niemi J., 2018. Good hygiene increases fattening pigs’ health and performance and benefits the farmer financially. In: PROHEALTH Sustainable pig and poultry production. Policy Briefs and Best Practice Guidelines, 16-17. https://www.fp7-prohealth.eu/wp-content/uploads/migrated/documents/PROHEALTH_Report-FINAL-Web.pdf
- Le Floc'h N., Otten W., Merlot E., 2011. Tryptophan metabolism, from nutrition to potential therapeutic applications. Amino Acids, 41, 1195-1205. https://doi.org/10.1007/s00726-010-0752-7
- Le Floc'h N., Gondret F., Resmond R., 2021. Identification of blood immune and metabolic indicators explaining the variability of growth of pigs under contrasted sanitary conditions. BMC Vet. Res., 17, 166. https://doi.org/10.1186/s12917-021-02872-3
- Liu Y., Espinosa C.D., Abelilla J.J., Casas G.A., Lagos V., Lee S.A., Kwon W.B., Mathai, J.K., Navarro D.M.D.L., Jaworski N.W., Stein H.H., 2018. Non-antibiotic feed additives in diets for pigs: A review. Anim. Nutr., 4, 113-125. https://doi.org/10.1016/j.aninu.2018.01.007
- Livestock Ontology, 2021. AHOL (Animal Health Ontology for Livestock) and ATOL (Animal Trait Ontology for Livestock). https://www.atol-ontology.com/
- Lundeheim N., 1987. Genetic Analysis of Osteochondrosis and Leg Weakness in the Swedish Pig Progeny Testing Scheme. Acta Agric. Scand., 37, 159-173. https://doi.org/10.1080/00015128709436552
- Madec F., Kobisch M., 1982. Bilan lésionnel des poumons des porcs charcutiers à l'abattoir. Journ. Rech. Porcine, 14, 405-412.
- Merlot E., Quesnel H., Prunier A., 2015. Conséquences du stress maternel pendant la gestation sur l'immunité et la santé des nouveau-nés en élevage. INRA Prod. Anim., 28, 283-294. https://doi.org/10.20870/productions-animales.2015.28.4.3033
- Merlot E., Calvar C., Prunier A., 2017. Influence of the housing environment during sow gestation on maternal health, and offspring immunity and survival. Anim. Prod. Sci., 57, 1751-1758. https://doi.org/10.1071/AN15480
- Merlot E., Pastorelli H., Prunier A., Père M.C., Louveau I., Lefaucheur L., Perruchot M.H., Meunier-Salaün M.C., Gardan-Salmon D., Gondret F., Quesnel H., 2019. Sow environment during gestation: Part I. Influence on maternal physiology and lacteal secretions in relation with neonatal survival. Animal, 13, 1432-1439. https://doi.org/10.1017/S1751731118002987
- Meunier-Salaün M.C., Guérin C., Billon Y., Sellier P., Noblet J., Gilbert H., 2014. Divergent selection for residual feed intake in group-housed growing pigs: characteristics of physical and behavioural activity according to line and sex. Animal, 8, 1898-1906. https://doi.org/10.1017/S1751731114001839
- Molist F., Van Oostrum M., Pérez J.F., Matéos G.G., C. Nyachoti M., Van Der Aar. P.J., 2014. Relevance of functional properties of dietary fibre in diets for weanling pigs. Anim. Feed. Sci. Technol., 189, 1-10. https://doi.org/10.1016/j.anifeedsci.2013.12.013
- Montagne L., Gilbert H., Muller N., Le Floc'h N., 2021. Physiological response to the weaning in two pig lines divergently selected for residual feed intake. J. Anim. Physiol. a. Anim. Nutr. First view. https://doi.org/10.1111/jpn.13622
- Murphy T.W., 2011. The effects of individual and combinations of airborne pollutants on feed intake, immune function and physiology of the pig. Thèse de doctorat. The University of Adelaide, Australia, 273p.
- Pastorelli H., Meunier-Salaün M.-C., Tallet C., Calvar C., Quesnel H., 2016. Effet de l'environnement des truies pendant la gestation sur leur comportement et la survie des porcelets. Journ. Rech. Porcine, 48, 201-206.
- Perrin W.R., Bowland J.P., 1977. Effects of enforced exercise on the incidence of leg weakness in growing boars. Can. J. Anim. Sci., 57, 245-253. https://doi.org/10.4141/cjas77-032
- Petersen J.S., Oksbjerg N., Jørgensen B., Sørensen M.T., 1998. Growth performance, carcass composition and leg weakness in pigs exposed to different levels of physical activity. Anim. Sci., 66, 725-732. https://doi.org/10.1017/S1357729800009309
- PROHEALTH consortium, 2015. Production diseases: The cost to pig producers. PROHEALTH Newsletter. https://www.fp7-prohealth.eu/knowledge-platform/newsletter-articles/production-diseases-the-cost/
- Prunier A., Heinonen M., Quesnel H., 2010. High physiological demands in intensively raised pigs: impact on health and welfare. Animal, 4, 886-898. https://doi.org/10.1017/S175173111000008X
- Quesnel H., Gondret F., Merlot E., Farmer C., 2015. Influences maternelles sur la consommation de colostrum et la survie néonatale du porcelet. INRA Prod. Anim., 28, 295-304. https://doi.org/10.20870/productions-animales.2015.28.4.3034
- Quesnel H., Peuteman B., Merlot E., Prunier A., Calvar C., Robert F., Meunier-Salaün M.C., 2018. Effet de l'enrichissement du milieu des truies pendant la gestation sur le stress maternel et la survie des porcelets. Journ. Rech. Porcine, 50, 275-280.
- Quesnel H., Père M.C., Louveau I., Lefaucheur L., Perruchot M.H., Prunier A., Pastorelli H., Meunier-Salaün M.C., Gardan-Salmon D., Merlot E., Gondret F., 2019a. Sow environment during gestation: Part II. Influence on piglet physiology and tissue maturity at birth. Animal, 13, 1440-1447. https://doi.org/10.1017/S1751731118003087
- Quesnel H., Peuteman B., Père M.C., Louveau I., Lefaucheur L., Perruchot M.H., Prunier A., Meunier-Salaün M.C., Gardan-Salmon D., Gondret F., Merlot E., 2019b. Effect of environmental enrichment with wood materials and straw pellets on the metabolic status of sows during gestation. Livest. Sci., 229, 43-48. https://doi.org/10.1016/j.livsci.2019.09.005
- Rhouma M., Fairbrother J.M., Beaudry F., Letellier A., 2017. Post weaning diarrhea in pigs: risk factors and non-colistin-based control strategies. Acta. Vet. Scand., 59, 31. https://doi.org/10.1186/s13028-017-0299-7
- Rutherford K.M.D., Donald R.D., Arnottt G., Rooke J.A., Dixon L., Mehers J.J.M., Turnbull J., Lawrence A.B., 2012. Farm animal welfare: assessing risks attributable to the prenatal environment. Anim. Welfare, 21, 419-429. https://doi.org/10.7120/09627286.21.3.419
- Schenck E.L., McMunn K.A., Rosenstein D.S., Stroshine R.L., Nielsen B.D., Richert B.T., Marchant-Forde J.N., Lay D.C., 2008. Exercising stall-housed gestating gilts: Effects on lameness, the musculo-skeletal system, production, and behavior. J. Anim. Sci., 86, 3166-3180. https://doi.org/10.2527/jas.2008-1046
- Sierźant K., Perruchot M.H., Merlot E., Le Floc'h N., Gondret F., 2019. Tissue-specific responses of antioxidant pathways to poor hygiene conditions in growing pigs divergently selected for feed efficiency. BMC Vet. Res., 15, 341. https://doi.org/10.1186/s12917-019-2107-2
- Stavrakakis, S. 2014. Biomechanical studies on locomotion in pigs. PhD Newcastle University. Faculty of Science, Agriculture and Engineering Newcastle upon Tyne, UK, 175p.
- Stavrakakis S., Loisel F., Sakkas P., Le Floc'h N., Kyriazakis I., Stewart G., Montagne L., 2019. A systematic literature mapping and meta-analysis of animal-based traits as indicators of production diseases in pigs. Animal, 13, 1508-1518. https://doi.org/10.1017/S1751731118002719
- van Grevenhof E.M., Ott, S., Hazeleger W., van Weeren P.R., Bijma, P., Kemp, B., 2011. The effects of housing system and feeding level on the joint-specific prevalence of osteochondrosis in fattening pigs. Livest. Sci., 135, 53-61. https://doi.org/10.1016/j.livsci.2010.06.010
- Ytrehus B., Carlson C.S., Ekman S., 2007. Etiology and Pathogenesis of Osteochondrosis. Vet. Pathol., 44, 429-448. https://doi.org/10.1354/vp.44-4-429
Résumé
Le projet européen PROHEALTH portait sur les maladies de production des porcs et des volailles. Pour l’espèce porcine, INRAE a apporté des connaissances nouvelles permettant de comprendre les mécanismes biologiques qui conduisent ou non à l’expression de ces maladies et de proposer des voies d’amélioration des conditions d’élevage pour préserver la santé des porcs. Ainsi, enrichir l’environnement des truies pendant la gestation a permis de réduire leur stress et donc d’améliorer leur bien-être et de réduire la mortalité néonatale des porcelets. Chez les porcelets, la dynamique de réponse aux stress du sevrage et la prévalence des problèmes digestifs ont été modulées par l’alimentation. Chez les porcs en croissance, augmenter la surface disponible en élevage contribuerait à prévenir l’apparition de troubles locomoteurs selon la prédisposition génétique des porcs à augmenter leur activité physique. Chez les porcs en croissance également, le respect de bonnes conditions d’hygiène du logement permet de préserver la santé et les performances de croissance en limitant l’inflammation systémique et les troubles respiratoires. En s’appuyant sur un modèle de lignées génétiques divergentes sélectionnées sur la Consommation Moyenne Journalière Résiduelle (CMJR), un indicateur de l’efficacité alimentaire, nous avons montré l’implication de facteurs génétiques dans la susceptibilité des porcs à certaines maladies comme les troubles digestifs du sevrage, l’ostéochondrose et l’inflammation causée par les mauvaises conditions d’hygiène du logement.
Pièces jointes
Pas de document complémentaire pour cet articleStatistiques de l'article
 Vues: 8009
Vues: 8009
Téléchargements
 XML: 326
XML: 326
 PDF: 923
PDF: 923
Articles les plus lus par le même auteur ou la même autrice
- Nathalie QUINIOU, Anne BOUDON, jean-Yves DOURMAD, Maud MOINECOURT, Nathalie PRIYMENKO, Agnès NARCY, Modélisation du besoin en calcium de la truie reproductrice et variation du rapport phosphocalcique des aliments selon le niveau de performance , INRAE Productions Animales: Vol. 34 No 1 (2021)
- Sophie PRACHE, Bénédicte LEBRET, Élisabeth BAÉZA, Bruno MARTIN, Joël GAUTRON, Françoise MÉDALE, Geneviève CORRAZE, Chloé VAN BAELEN, Mégane RAULET, Florence LEFÈVRE, Véronique VERREZ-BAGNIS, Pierre SANS, Qualité et authentification des produits animaux issus de l’agriculture biologique vs conventionnelle , INRAE Productions Animales: Vol. 37 No 2 (2024): Numéro spécial : L’élevage biologique : conditions et potentiel de développement
- Florence GONDRET, Donato ANDUEZA, Mohamed Habibou ASSOUMA, Valérie BERTHELOT, Didier BOICHARD, Maguy EUGÈNE, Solène FRESCO, Amandine LURETTE, Cécile MARTIN, Pauline MARTIN, Diego P. MORGAVI, Rafael MUÑOZ-TAMAYO, Milka POPOVA, Simon ROQUES, Flavie TORTEREAU, Xavier FERNANDEZ, Réduction des émissions de méthane entérique chez les ruminants : enjeux, solutions et perspectives à l’échelle de l’animal et des systèmes d’élevage , INRAE Productions Animales: Vol. 39 No 1 (2026)
- Pierre NOZIÈRE, René BAUMONT, Anne BOUDON, Rémy DELAGARDE, Philippe HASSOUN, Sophie LEMOSQUET, Gaëlle MAXIN, Bernard SEPCHAT, Luc DELABY, Le système d’alimentation INRA 2018 pour les ruminants. 2. Application au rationnement , INRAE Productions Animales: Vol. 39 No 2 (2026)
- Pierre NOZIÈRE, René BAUMONT, Anne BOUDON, Rémy DELAGARDE, Philippe HASSOUN, Sophie LEMOSQUET, Gaëlle MAXIN, Bernard SEPCHAT, Luc DELABY, Le système d’alimentation INRA 2018 pour les ruminants. 1. Bases conceptuelles , INRAE Productions Animales: Vol. 39 No 2 (2026)